 |
| Reconstrucción por ordenador de los cinco cráneos de Dmanisi. Imagen: Marcia Ponce de León y Christoph Zollikofer de la Universidad de Zúrich |
Homo georgicus, Gabunia et al, 2002.
 |
Comparación de varios ejemplares de erectus y georgicus.
|
- La calota completa D 2280. Conserva parte de la región basal del cráneo con las partes basal y nucal del occipital ligeramente dañadas, partes del ala grande del esfenoides y la mayoría de la fosa mandibular temporal.
- Giorgy Nioradze halló el cráneo D 2282, calota y fragmento de maxilar. Deformado. Los maxilares retienen piezas dentales ligeramente desgastadas y alveolos de dientes que son visibles en radiografía. Posiblemente perteneció a una hembra joven.
 |
| Mandíbulas de Dmanisi. D 211, D 2600 y D 2735. |
 |
| D 211 y D 2600 presentan polaridades molares opuestas |
En 2000 se extrajo la mandíbula D 2600. Solo le falta el
extremo inferior de las ramas mandibulares. Muy grande, la mayor atribuida al
género Homo; por esta razón se ha
especulado con su pertenencia a otra especie diferente. Ramas mandibulares muy
altas. Incisivos de tamaño muy pequeño, especialmente los centrales. Caninos
grandes comparativamente. Premolares con dos raíces, lo cual es un carácter muy
arcaico. Molares grandes y masivos, de esmalte grueso. Polaridad opuesta a la
de D 211: tercer molar más grande que el segundo y este más que el primero.
Buena parte de estas características coinciden con las de habilis y rudolfensis,
pero el tamaño es mayor. Por ello, Leo Gabunia propuso la nueva especie Homo georgicus.
- Para Skinner et al (2008) las diferencias entre D 2600 por una parte, y D 211 y D 2735 por la otra, son demasiado grandes para que se deban a dimorfismo sexual. José María Bermúdez de Castro ha apuntado que en este caso de hipotético dimorfismo sexual se daría la paradoja de que el individuo más grande, “el macho”, tendría una capacidad craneal mucho más pequeña que las supuestas hembras.
- Para Ann Margvelashvili et al (2013), usando las poblaciones modernas de cazadores-recolectores como referencia, las variaciones en características como la forma de la arcada dental, la altura del corpus mandibular o la inclinación de la sínfisis, permanecen dentro del rango normal. El desgaste excesivo de los dientes conduce a la ruptura de los mecanismos normales de remodelación, lo que resulta en patologías, pérdida de dientes y pérdida de la función masticatoria y además en la extraordinaria altura de la mandíbula. Sin embargo, es poco probable que ello haya limitado la duración de la vida porque este efecto fue probablemente paliado por la preparación de alimentos blandos.
- María Martinón-Torres (2013) señala la posibilidad de que esta mandíbula no sea patológica. En todo caso, las patologías habrían producido una recesión del borde superior de la mandíbula. Además resalta que del análisis del desgaste se deriva una alimentación herbívora, similar a la de chimpancés y gorilas. Esta dieta sustancialmente vegetal podría sugerir una adaptación a un nicho ecológico diferente para el individuo del cráneo 5 y los demás homínidos encontrados en el yacimiento. Además explicaría la ausencia de mecanismos compensatorios (como la migración dental mesial o mesial drift) habituales en Homo. En las mandíbulas D 211 y D 2375 se identifican estos cambios compensatorios, lo que evidencia un uso sustancialmente diferente de los dientes entre ambos conjuntos.
- Laura Martín-Francés et al (2014) detectan un desgaste dental severo e inusual, acompañado de una extensa exposición de la raíz con migración axial, abscesos periapicales y fracturas en el esmalte. Además, hay evidencia de rotación dental post-eruptiva y artropatía temporomandibular. Proponen que el patrón de desgaste observado en este individuo, similar al registrado en chimpancés y gorilas y diferente al observado en el resto de Homo, se relaciona con una dieta con un alto consumo de alimentos fibrosos y abrasivos tales como frutas y vegetales. Las superficies oclusales redondeadas y las superficies labio-linguales muy pulidas de los dientes anteriores de D 2600 podrían ser principalmente la consecuencia de actividades pre o para-masticatorias, como la sujeción y la extracción. Este tipo de comida sería también el origen de la morfología oclusal en forma de copa de la dentición posterior en combinación con un relativamente ligero desgaste proximal. Las lesiones mostradas por D 2600 no han alterado significativamente la morfología de la mandíbula y no impiden una evaluación taxonómica apropiada.
- Según Ann Margvelashvili et al (2016), las patologías son más prevalentes y el desgaste está más avanzado en las muestras de Dmanisi que en los actuales cazadores recolectores.
- Para José María Bermúdez de Castro et al (2014), la combinación de rasgos arcaicos y derivados en D 2600 puede estar relacionada con la dieta. Las principales características derivadas de D 2600 con respecto a las otras mandíbulas, son las siguientes:
- Presencia de un área retromolar subhorizontal y bien desarrollada.
- Alineación de corpus y ramus, con valores similares a los ejemplares de Homo ergaster (Rosas A, Bermúdez de Castro JM; 1998).
- Línea milohioidea visible e inclinada. Esta es una característica típica del linaje neandertal.
- Prominencia lateral débil al nivel de M2-M3.
- Fosa masetérica superficial.
- Borde anterior del ramus desplazado hacia delante, cubriendo solo parte de M3.
- Jeffrey H. Schwartz, Ian Tattersall y Zhang Chi (2014) destacan asimismo las diferencias entre D 2600 por un lado y D 2735 y D 211 por otro.
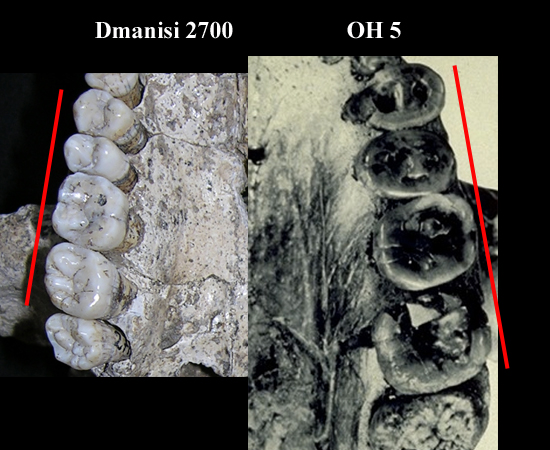 |
| D 2700 Homo georgicus y OH 5 Paranthropus boisei |
 |
| TM 1517, Paranthropus robustus y D 2735 Homo georgicus |
En 2001 apareció el cráneo D 2700 en extraordinario estado de conservación y la mandíbula D 2735, completa, que encaja en el cráneo anterior, que pertenecieron a un macho subadulto. Cráneo pequeño, dentro del rango de habilis. Cara baja y cóncava, de caninos prominentes y acusado prognatismo. Grandes caninos. Incisivos grandes y en forma de paleta. Caninos grandes y prominentes.
 |
| Sts 5, Australopithecus africanus y D 3444, Homo georgicus |
En 2002 Slava Ediberidze encontró el cráneo D 3444 de 650 cc, y la mandíbula D 3900, asociados a industria olduvaiense.
En 2003 se halló una mandíbula que recibió el nombre de Viejo de Dmanisi, ya que perteneció a un
anciano (40-50 años) que había perdido todos sus dientes hacía mucho tiempo,
por lo que debió de haber sido alimentado por otros humanos.
En 2005 se halló el Cráneo 5 (D 4500; David Lordkipanidze et al, 2013; Rightmire et al, 2017) que ha resultado corresponder al mismo individuo que la mandíbula D 2600. Esta combinación mezcla de forma inaudita un cerebro pequeño (540 cc) con una cara muy prognática y exhibe afinidades morfológicas con los Homo tempranos de África. Este individuo sería un varón que medía entre 1,40 y 1,60 m y pesaba alrededor de 50 kg. Cuando murió debía tener unos 30 años.
En 2005 se halló el Cráneo 5 (D 4500; David Lordkipanidze et al, 2013; Rightmire et al, 2017) que ha resultado corresponder al mismo individuo que la mandíbula D 2600. Esta combinación mezcla de forma inaudita un cerebro pequeño (540 cc) con una cara muy prognática y exhibe afinidades morfológicas con los Homo tempranos de África. Este individuo sería un varón que medía entre 1,40 y 1,60 m y pesaba alrededor de 50 kg. Cuando murió debía tener unos 30 años.
En 2011 se encontró un hueso de dedo del pie.
Además se han hallado un fémur, un peroné, varias tibias,
vértebras, costillas y diversos huesos de la mano y el pie. Las proporciones
entre los brazos y las piernas son claramente modernas. El pie muestra un arco
bien desarrollado, con el dedo gordo en posición paralela. El peso máximo sería
de 50 kg y la altura máxima 1,50 m. Las palmas de las manos estaban orientadas
hacia delante, lo que se interpreta como una adaptación a la vida arbórea.
 |
Vistas de los cráneos de Dmanisi. Escala 5 cm. De arriba a abajo:
D2700/D2735, adolescente.
D2282/D211, adulto joven (reconstrucción virtual).
D2280, adulto.
D4500/D2600, adulto.
D3444/D3900 adulto anciano edéntulo.
David Lordkipanidze et al, 2013
|
Según Philip Rightmire y David Lordkipanidze (2006) los caracteres identificativos de estos fósiles son:
- Plesiomorfias (caracteres primitivos)
- Baja capacidad craneal (600-775 cc).
- Pronunciado estrechamiento frontal (índice de constricción postorbital 68,7-74,4).
- Región mastoidea inflada y proyectada lateralmente.
- Toro transversal occipital pobremente expresado o ausente.
- Índice occipital bajo: 85-102,1.
- Proyección del contorno del tercio medio facial.
- Jugum canino prominente y limitado lateralmente por el surco maxilar
- Incisura cigomáticoalveolar presente.
- Barbilla ligera o ausente.
- Sínfisis reforzada internamente (toro tansversal superior).
- Sinapomorofias con Homo erectus (senso lato).
- Toro supraorbital en forma de barra y saliente.
- Fontanela anterior con quilla sagital parietal.
- Toro angular presente.
- Escama del temporal baja, con el borde superior recto.
- Punta de la mastoides envuelta y aplanada posteriormente.
- Ángulo petrotimpánico bajo (140º-150º).
- Pirámide petrosa lisa, densa en apariencia.
- Foramen lacerum restringido.
- Silla nasal prominente.
- Posibles sinapomorfías con Homo erectus asiáticos.
- Aplanamiento parasagital (depresión) de las superficies parietales.
- Crestas paramastoidea y occipitomastoidea presentes.
- Posibles autapomorfías.
- Doble quilla sagital.
- Placa timpánica relativamente delicada.
- Proceso supratubario de Weindenreich ausente.
- M3 es de tamaño reducido.
 |
| SK 847 (arriba), D4500 (medio) y AL 666-1 (abajo). Las flechas señalan similitudes. Escala 5 cm. David Lordkipanidze et al, 2013 |
Interpretaciones filogenéticas
La presencia de estos restos en Georgia nos indica que la primera salida homínida de África, corrió a cargo de una especie de cerebro pequeño, próxima a H. habilis.
Para Emiliano Aguirre (2008) los seres de Dmanisi
pertenecen a un grupo que salió de África en el momento de la cladogénesis del ergaster.
| Parte distal del corpus de D 2735 (izquierda) y D 2600 (derecha) |
En 2008 su equipo analizó al detalle la dentición de los homínidos de Dmanisi, destacando el aspecto marcadamente primitivo de algunos de los dientes georgianos, en el caso de los caninos, semejantes incluso a especímenes de H. habilis o incluso del género Australopithecus. Los dientes de D 2600 tienen las coronas muy gastadas y su longitud, número y grado de bifurcación y molarización es marcadamente robusto y primitivo y, en su conjunto, solo presente en algunos especímenes de Homo rudolfensis, Australopithecus o incluso Paranthropus. En las otras dos mandíbulas, en D 211 y D 2375, a pesar de los rasgos habilinos, se dan unas características sorprendentemente derivadas (o “modernas”) para una población de esta antigüedad. Los dientes de D 211 y D 2375 presentan una serie molar decreciente (el primer molar es mayor que el segundo que es a su vez mayor que el tercero) y otras características relacionadas con un tamaño dental reducido que son inusuales para una población de esa antigüedad. Sugieren la posibilidad de que en el hipodigma de Dmanisi hubiera representadas al menos dos paleodemes o poblaciones. De la misma opinión son Jeffrey H. Schwartz, Ian Tattersall y Zhang Chi (2014).
Para Pontzer et al (2011), los molares de los seres de Dmanisi son similares a los del ergaster. La dentadura no estaba especializada lo que indica que georgicus explotaba una amplia gama de alimentos.
Para Pontzer et al (2011), los molares de los seres de Dmanisi son similares a los del ergaster. La dentadura no estaba especializada lo que indica que georgicus explotaba una amplia gama de alimentos.
David Lordkipanidze et al (2013) suponen que los cinco cráneos hallados en Dmanisi se depositaron en un intervalo de unos 20 ka y pertenecen a un mismo demo muy variable. Ello daría pie a reconocer un rango muy amplio de variación morfológica en los primeros Homo y por consiguiente reunirlos en un único linaje evolutivo (habilis, rudolfensis, ergaster y erectus) (Rightmare et al, 2017).
- Adam van Arsdale y Milford H. Wolpoff (2012) son partidarios asimismo de un único linaje.
 |
| D4500 y D2600 comparten características con todas las especies Homo tempranas, lo que sugiere un único linaje evolutivo muy variable. |
- Esta interpretación está siendo muy discutida. Parece difícil que no se haya producido especiación en demos muy reducidas, aisladas durante centenares de miles de años y explotando entornos muy diferentes. José María Bermúdez de Castro (2013) apunta que D 4500+D 2600 representa el cráneo más pequeño de la muestra al lado del aparato masticador más robusto, lo que induce a pensar que pertenece a otra especie diferente.
- Si la mandíbula D 2600 no es patológica se hace difícil de explicar su relación con el resto de la muestra.
| Ubicación estratigráfica de los hallazgos de Dmanisi |
- Calvo-Rathert (2008) sugieren que la acumulación de los homínidos de Dmanisi podría cubrir un periodo de hasta cientos de miles de años. La mandíbula D2 600 proviene de una capa con polaridad normal (es decir, pertenece al subcrón Olduvai y por lo tanto tiene más de 1,78 Ma) y algunos de los demás fósiles se han encontrado en capas con polaridad magnética reversa (post-Olduvai y por lo tanto más jóvenes que 1,78 Ma) (María Martinón-Torres, 2013).
- Los fósiles se hallaron principalmente en dos áreas diferentes. Bloque 1 (D 211, D 2280, D 2282) y Bloque 2 (D 2735, D 2700, D 4500 y D 2600) separados de 10 a 20 m. Varios fósiles (D 2280, D 2282, D 211, D 2700 y D 2735) se atribuyeron a la unidad estratigráfica A, de polaridad normal (subcrón Olduvai), pero más tarde Ferring et al comunicaron que todos los fósiles humanos habían sido recuperados de los estratos inversamente polarizados B1x-B1z.
- Un metatarsiano de adulto se halló en B1z
- D 2700 y D 2735, se asignan a B1x.
- D 2600, D 4500, D 3900 y D 3444 se asignan a la unidad B1y.
- D 2280, D 2282 y D 211 no han sido atribuidos de forma específica a ninguna unidad estratigráfica.
- Varios investigadores, como Lee Berger o María Martinón-Torres, han destacado la necesidad de comparar los ejemplares de Daminisi con Paranthropus y Australopithecus.
- José María Bermúdez de Castro et al (2014) apoyan la existencia de dos paleodemos diferentes en Dmanisi: D 211 y D 2735 estarían viuculados a Homo habilis o Homo ergaster, mientras que las características en mosaico de D 2600 no se encuentran en ningún otro homínido y justifican la independencia del taxón Homo georgicus.
Herramientas líticas
El 82,7% corresponde a bases positivas o lascas de los que solo un 4,5% sobre el total corresponde a objetos retocados de pequeño formato. Un 33,2% de las lascas son corticales. Los núcleos representan un 4,3% del total y los choppers y chopping-tools un 10,4%.
Los análisis petrográficos demuestran que hubo traslado de ciertos materiales hasta el propio yacimiento, lo cual indica una cierta complejidad en el proceso de adquisisicón de las materias primas. La talla está completamente condicionada por la geometría natural de los soportes, a menudo ortogonal con ángulos de percusión muy altos, si bien se presenta una cierta diversidad de sistemas de talla. Los núcleos, lascas y debrises presentes, demuestran además que todo el proceso de reducción se elaboró en el mismo sitio. Numerosos núcleos unifaciales sugieren que no se trataba de una talla “demasiado elaborada”. También se documenta talla centrípeta en algunos núcleos sobre lasca (flake-cores). Sin bifaces. Predominio absoluto de lascas frente a herramientas. El sitio es comparable a Bed I de Olduvai o a Atapuerca TD-6.
El clima de Dmanisi, por entonces, debió ser moderadamente árido, con altas montañas surcadas por profundos valles y cuadalosos ríos, con paisajes abiertos de estepa cálida en las llanuras circundantes surcadas por bosques-galería en los valles. Por consiguiente, los homínidos de Dmanisi tuvieron a su disposición una considerable diversidad de hábitats.
Javier Baena ha identificado dos distintos conjuntos culturales.
Un hueso de venado hallado junto al cráneo 5 conservaba una escama lítica incrustada; marcas de herramientas en algunos otros huesos de animales, sugieren el consumo humano. Pero, por otra parte, los carnívoros parecen ser los agentes primarios de las acumulaciones fósiles.
| Mandíbulas de Dmanisi. La flecha de arriba a la izquierda señala una lesión producida por el uso continuado de palillo. |
La salida de África.
La misma crisis climática que condujo a la aparición del ergaster obligó al habilis y a otros seres parecidos a migrar siguiendo las masas de
bosque. Algunas poblaciones llegaron probablemente al Centro de África, donde
todavía hoy se mantienen los bosques tropicales. El bosque tropical es poco
propicio para la conservación de fósiles. Otras poblaciones se dirigieron hacia
el Norte, siguiendo las riberas del Nilo y el Mar Rojo. En el Cáucaso
encontraron un hábitat idóneo.
En 2015, Wei Qi comunicó en nota de prensa una datación de 1,95-1,77 Ma para el yacimiento de Heitugou, cuenca del Nihewan, Yangyuan, China, que sería por tanto anterior a Dmanisi.
En 2015, Wei Qi comunicó en nota de prensa una datación de 1,95-1,77 Ma para el yacimiento de Heitugou, cuenca del Nihewan, Yangyuan, China, que sería por tanto anterior a Dmanisi.
| Detalle de la parte izquierda de D 2600, donde se precibe una raiz molarizada bifurcada en P3 |
Extracción del cráneo 5.
Adam van Arsdale: Homo georgicus.
Bibliografía:
- Gabunia, Leo, et al. "Dmanisi and dispersal." Evolutionary Anthropology: Issues, News, and Reviews 10.5 (2001): 158-170.
- Vekua, Abesalom, et al. "A new skull of early Homo from Dmanisi, Georgia."Science 297.5578 (2002): 85-89.
- Rightmire, G. Philip, David Lordkipanidze, and Abesalom Vekua. "Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia." Journal of Human Evolution 50.2 (2006): 115-141.
- Lordkipanidze, David, et al. "Postcranial evidence from early Homo from Dmanisi, Georgia."Nature 449.7160 (2007): 305-310.
- Lordkipanidze, David, et al. "A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo." Science 342.6156 (2013): 326-331.
- Bermúdez de Castro JM, Martinón-Torres M, Sier MJ, Martín-Francés L (2014) On the Variability of the Dmanisi Mandibles. PLoS ONE 9(2): e88212. doi:10.1371/journal.pone.0088212
Te puede interesar:
|
No hay comentarios:
Publicar un comentario