| Localizacion de Malapa. Aurore Val et al, 2015. |
 |
| MH1 (Karabo). Australopithecus sediba. Lee Berger, University of Witwatersrand. |
 |
| Cráneo de Australopithecus sediba. Reconstrucción por Peter Schmid, foto de Lee R. Berger. |
 |
| Australopithecus sediba. Recreación de John Gurche. |
Australopithecus sediba, Berger et al 2010
Fue hallado por Matthew Berger, hijo de Lee R. Berger, de la universidad de
Witwatersrand, Johannesburg, Sudáfrica, el 15 de agosto de 2008 en Malapa,
Sudáfrica, a 15 km. de Sterkfontein, Swartkrans, y Kromdraai.
El yacimiento de Malapa se encuentra 15 km al nordeste de Sterkfontein. Los restos de Australopithecus sediba (MH1 y MH2), aparecieron al excavar un pozo de 3,3 m x 4,4 m y 4 m de profundidad, con sedimentos resultado de la erosión de un sistema antiguo de cuevas profundas, en la Facies D, datada en 1,977 ± 0.002 Ma, formada por un flujo de escombros, mientras que los sedimentos suprayacentes pertenecientes a las facies E y F se depositaron en forma de acumulaciones de barro a lo largo del suelo de la cueva, con la afluencia ocasional de material arenoso, como resultado de una corriente de agua. Durante gran parte de su historia deposicional, los sedimentos de las facies E y F fueron anegados. En términos de composición, los sedimentos de las facies D, E y F, representan una mezcla de depósitos mal ordenados autóctonos y alóctonos que contienen una alta proporción de material peloidal rico en manganeso, derivado de la reelaboración de sedimentos fangosos por insectos. En general, los sedimentos van haciéndose más finos, con arenas gruesas con bloques en la base de la facies E, hasta barro puro en la parte superior de la facies F, con bloques de dolomita aislados y fósiles incrustados. Según el estudio tafonómico de Aurore Val et al (2015) y Brophy et al (2016), la diversidad del espectro de fauna, la presencia de animales con inclinaciones de escalada, la abundancia de especímenes completos y/o articulados y la falta de huesos modificados por carnívoros, indican que los animales se acumularon a través de una trampa en el sistema de cuevas sin acceso a los carroñeros. La co-ocurrencia de fósiles bien conservados, coprolitos de carnívoros, dientes de leche de hiena marrón, y algunos restos muy fragmentados y mal conservados apoya la hipótesis de una mezcla de sedimentos procedentes de cámaras distintas, recogidos en el fondo del sistema de cuevas por la acción de las corrientes de agua. Esta combinación de características tafonómicas explica el notable estado de conservación de los fósiles de Hominini, así como parte del material de la fauna asociada. Los restos no están asociados a herramientas.
Ericka N. L’Abbé et al (2015) han evaluado todas las fracturas evidentes en MH1 y MH2. La mayor parte de las fracturas son post-deposicionales, pero las del miembro superior derecho de la hembra adulta junto con posibles fracturas en la mano, indican claramente la resistencia activa a un impacto, mientras que las de la mandíbula del juvenil son consistentes con un golpe en la cara. La presencia de estos traumatismos apoya la hipótesis de la caída y suministra la primera evidencia de muerte de un australopitecino por causas diferentes a la depredación o la muerte natural, si bien en el caso de MH1 no se puede descartar la violencia interpersonal. Los Hominini de Malapa pudieron haber caído en un hoyo en la piedra caliza, posiblemente entre 5-10 m de profundidad. La superficie de impacto estuvo formada probablemente por grava, arena y guano de murciélago, con una moderada a alta capacidad de absorción de energía. Si bien las fracturas perimortem observadas en MH2 no son fatales por sí mismas, no se observan signos macroscópicos de curación, por lo que la muerte llegó poco después de las lesiones. Después de la deposición, un entierro natural rápido impidió el acceso de los carnívoros y la dispersión de los restos lo que añadido a un pH favorable, puede explicar la excelente conservación. La ubicación de MH1 y MH2 sugiere que entraron en la cueva y murieron en un lapso temporal breve. Posteriormente, el techo de la cámara se derrumbó y se sucedieron repetidas caídas de rocas. Más tarde, la cueva se llenó de barro y de otros materiales paleontológicos.
El yacimiento de Malapa se encuentra 15 km al nordeste de Sterkfontein. Los restos de Australopithecus sediba (MH1 y MH2), aparecieron al excavar un pozo de 3,3 m x 4,4 m y 4 m de profundidad, con sedimentos resultado de la erosión de un sistema antiguo de cuevas profundas, en la Facies D, datada en 1,977 ± 0.002 Ma, formada por un flujo de escombros, mientras que los sedimentos suprayacentes pertenecientes a las facies E y F se depositaron en forma de acumulaciones de barro a lo largo del suelo de la cueva, con la afluencia ocasional de material arenoso, como resultado de una corriente de agua. Durante gran parte de su historia deposicional, los sedimentos de las facies E y F fueron anegados. En términos de composición, los sedimentos de las facies D, E y F, representan una mezcla de depósitos mal ordenados autóctonos y alóctonos que contienen una alta proporción de material peloidal rico en manganeso, derivado de la reelaboración de sedimentos fangosos por insectos. En general, los sedimentos van haciéndose más finos, con arenas gruesas con bloques en la base de la facies E, hasta barro puro en la parte superior de la facies F, con bloques de dolomita aislados y fósiles incrustados. Según el estudio tafonómico de Aurore Val et al (2015) y Brophy et al (2016), la diversidad del espectro de fauna, la presencia de animales con inclinaciones de escalada, la abundancia de especímenes completos y/o articulados y la falta de huesos modificados por carnívoros, indican que los animales se acumularon a través de una trampa en el sistema de cuevas sin acceso a los carroñeros. La co-ocurrencia de fósiles bien conservados, coprolitos de carnívoros, dientes de leche de hiena marrón, y algunos restos muy fragmentados y mal conservados apoya la hipótesis de una mezcla de sedimentos procedentes de cámaras distintas, recogidos en el fondo del sistema de cuevas por la acción de las corrientes de agua. Esta combinación de características tafonómicas explica el notable estado de conservación de los fósiles de Hominini, así como parte del material de la fauna asociada. Los restos no están asociados a herramientas.
Ericka N. L’Abbé et al (2015) han evaluado todas las fracturas evidentes en MH1 y MH2. La mayor parte de las fracturas son post-deposicionales, pero las del miembro superior derecho de la hembra adulta junto con posibles fracturas en la mano, indican claramente la resistencia activa a un impacto, mientras que las de la mandíbula del juvenil son consistentes con un golpe en la cara. La presencia de estos traumatismos apoya la hipótesis de la caída y suministra la primera evidencia de muerte de un australopitecino por causas diferentes a la depredación o la muerte natural, si bien en el caso de MH1 no se puede descartar la violencia interpersonal. Los Hominini de Malapa pudieron haber caído en un hoyo en la piedra caliza, posiblemente entre 5-10 m de profundidad. La superficie de impacto estuvo formada probablemente por grava, arena y guano de murciélago, con una moderada a alta capacidad de absorción de energía. Si bien las fracturas perimortem observadas en MH2 no son fatales por sí mismas, no se observan signos macroscópicos de curación, por lo que la muerte llegó poco después de las lesiones. Después de la deposición, un entierro natural rápido impidió el acceso de los carnívoros y la dispersión de los restos lo que añadido a un pH favorable, puede explicar la excelente conservación. La ubicación de MH1 y MH2 sugiere que entraron en la cueva y murieron en un lapso temporal breve. Posteriormente, el techo de la cámara se derrumbó y se sucedieron repetidas caídas de rocas. Más tarde, la cueva se llenó de barro y de otros materiales paleontológicos.
Lee R. Berger y otros, estudiaron los restos de Hominini,
pertenecientes a una hembra adulta y a un joven (Karabo = La Respuesta), sin poder encuadrarlos en taxones
existentes. Por ello, establecieron una nueva especie, Australopithecus sediba. Sediba
significa manantial inagotable en la lengua seSotho. Las diferencias entre ambos especímenes indican un amplio rango de variación en la especie (Ritzman et al, 2016).
Comparte con Australopithecus africanus
numerosas características en la bóveda del cráneo, el esqueleto facial, la
mandíbula y los dientes. Pero otras características (dientes más pequeños,
pómulos menos pronunciados, nariz prominente, piernas más largas, rasgos de la
pelvis) lo acercan a Homo. No
obstante, en base a algunos rasgos entre los que destaca el pequeño tamaño del
cerebro, se ha incluido dentro del género Australopithecus.
 |
| Australopithecus sediba. Dientes derechos maxilares (izquierda) y mandibulares (derecha). |
 |
| UW88-50 (MH1) elementos craneodentales en A. Vista superior; B. Vista frontal; C. Vistas laterales y D. mandíbula juvenil UW88-8 (MH1). (Berger, et al. 2010). |
 |
| Comparación de los coxis de Au. afarensis (AL 288-1), Au. africanus (Sts 14), Au. sediba (MH1), y H. erectus (KNM-WT 15000). Science. |
 |
| Australopithecus sediba. MH1 y MH2. Science. |
 |
| Australopithecus sediba. Reconstrucción utilizando MH1 y MH2. Lee R. Berger. |
El paratipo es MH2, una hembra adulta representada por un
diente maxilar, una mandíbula parcial, y parte del esqueleto postcraneal. Del
análisis de las manos, con dedos cortos y pulgar relativamente largo, capaces
de realizar la pinza de precisión, puede inferirse la capacidad para producir
herramientas líticas.
Los largos brazos y la estructura de los pies sugieren
capacidad de trepa. Se deduce que esta especie disfrutaba de una amplia
variedad de comportamientos y podía explotar diversos entornos.
Se estima que la altura máxima de la especie era 1,56 m
En MH1 y MH2 se han encontrado restos de tejidos blandos.
Fueron encontrados otros restos, correspondientes al menos a
un bebé y a una hembra adulta, pero aún no se han publicado los resultados.
Ella Been y Yoel Rak piensan que MH1 y MH2 son en realidad una mezcla de restos de Homo y Australopithecus.
Ella Been y Yoel Rak piensan que MH1 y MH2 son en realidad una mezcla de restos de Homo y Australopithecus.
Los estudios confirman una evolución en mosaico:
- Rasgos primitivos.
- Tórax superior estrecho y cónico. Esta morfología limita el balanceo de los brazos al caminar. La forma de las costillas es más similar a la de los chimpancés que la la de los humanos (Bastir et al, 2016). Las vértebras cervicales indican que el cuello era más rígido que en Homo, una mayor elevación del brazo consistente con comportamientos de trepa y la ausencia de un control motor mejorado en los miembros superiores (Meyer et al, 2017).
- Brazos largos y simiescos y escápula adaptados a la trepa e incluso la suspensión (Rein et al, 2016).
- Pie similar al del chimpancé, aunque con un tobillo como el de Homo. El talón es todavía más primitivo que el de Australopithecus afarensis. Según Thomas C. Prang (2015) A. sediba es indistinguible de los simios africanos existentes en la configuración angular de su parte trasera del pie, lo que sugiere que carecía de arco longitudinal y combina de forma única la morfología articular talocalcaneal, responsable de la movilidad, con especializaciones de la articulación talonavicular que proporcionan estabilización mediotarsiana medial. A. sediba es muy similar a los gorilas existentes en la morfología de la articulación subastragalina. La similitud morfológica entre MH2 y las muestras de Sterkfontein, miembro 4 (STW 88, STW 102, STW 352) en las articulaciones talonavicular y talocalcaneal sugiere que A. sediba puede haber poseído un pie que era funcionalmente similar al de A. africanus. Esta combinación de morfologías sugiere que arboralidad puede haber sido significativamente adaptativa para los Australopithecus del sur de África.
- Rasgos derivados en la línea del sapiens.
- Curvatura baja de la espalda. Columna flexible. Mismo número de vértebras lumbares que Homo. Lordosis cervical, mitigación de la carga y soporte de la cabeza similares a Homo (Meyer et al, 2017).
- Pelvis
- Muñeca, mano y dedos adaptados al agarre de precisión. Pudieron fabricar herramientas sin perder la capacidad de escalada. Aunque no se hallaron herramientas líticas en asociación directa, la fabricación de herramientas (de piedra y hueso) se ha documentado en otros yacimientos contemporáneos locales, como Sterkfontein y Swartkrans. Los huesos de la mano presentan un mosaico único de características. Las falanges proximales e intermedias son ligeramente curvadas, como en Au. africanus y OH 7 y muestra un desarrollo de la envoltura sinovial del flexor que indica un poderoso agarre. Los huesos de la muñeca también muestran más similitudes con otros australopitecinos que con Homo y los ejes metacarpianos, sobre todo el del primer metacarpiano, son notablemente gráciles (aunque el tercer metacarpiano del juvenil masculino MH1 es más robusto, lo que sugiere un fuerte grado de dimorfismo sexual en la morfología mano). El pulgar es largo en relación con los otros dedos y relativamente más largo que el de los HAM, cayendo fuera del rango de variación. La longitud del pulgar pudo permitir un mayor control para la manipulación de los objetos pequeños (Kivell, 2015). La morfología trabecular de los metacarpianos es similar a la de los orangutanes, consistente con el uso locomotor con los dedos, mientras que la del pulgar es consistente con la manipulación humana (Dunmore et al, 2020).
- Dientes.
- Boca parecida a la humana.
- Caninos pequeños.
- Aunque relacionados, son distintos a los de Australopithecus africanus en tamaño, forma y trayectoria ontogenética y esto garantiza la separación filogenética con esta especie.
- Los dientes de A. sediba (y tambien los de A. africanus) están más relacionados con los de Homo que los dientes de los austalopitecinos del Este de África (Irish et al, 2016).
- Sugieren que Australopithecus sediba no desciende de Australopithecus afarensis.
- Cuatro de los rasgos compartidos por Au. sediba, Au. africanus y los Homo tempranos no se ven en los primeros homínidos.
- El cerebro, aunque es pequeño, posee una región frontal expandida lo que indica una avanzada reorganización de la materia gris.
- Otros rasgos derivados
- Caminaba con hiperpronación (con rotación hacia dentro de rodilla y cadera y con los pies ligeramente torcidos hacia dentro) y pasos cortos y rápidos, apoyando primero el borde exterior del pie en lugar del talón.
- La mandíbula de MH1 presenta rasgos idénticos a otros australopitecinos, pero la mandíbula de MH2 es similar a la de Homo. Por el contrario, las dos vértebras lumbares de MH1, son similares a las de Homo, mientras que las de MH2 se parecen a las de los australopitecinos. Esto se ha interpretado como una gran variabilidad dentro de la especie.
 |
| Australopithecus sediba. Restos correspondientes a las extremidades superiores. |
 |
| Características del Australopithecus sediba. |
MH1 y MH2 se han datado en 1,977 Ma. por lo que representan
a los Australopithecus más modernos.
En 2012 se notificó el hallazgo de un esqueleto muy completo
de un joven, que podría corresponder con MH1 Karabo.
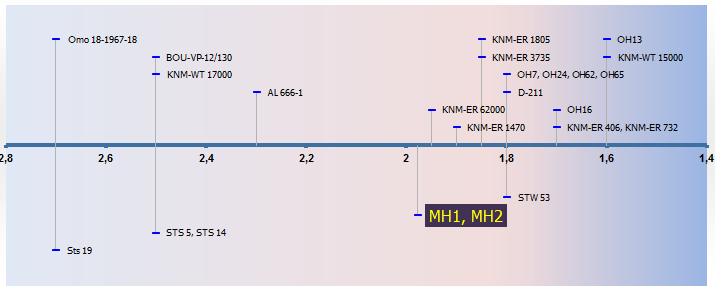 |
| Ubicación cronológica de Australopithecus sediba |
| MA | Ejemplares | Especie |
| 2,7000 | Sts 19 | Homo? |
| 2,7000 | Omo 18-1967-18 | Paranthropus aethiopicus |
| 2,5000 | BOU-VP-12/130 | Australopithecus garhi |
| 2,5000 | STS 5, STS 14 | Australopithecus africanus |
| 2,5000 | KNM-WT 17000 | Paranthropus aethiopicus |
| 2,3000 | AL 666-1 | Homo? |
| 1,9770 | MH1, MH2 | Australopithecus sediba |
| 1,9500 | KNM-ER 62000 | Homo rudolfensis |
| 1,9000 | KNM-ER 1470 | Homo rudolfensis |
| 1,8500 | KNM-ER 1805 | Homo rudolfensis |
| 1,8500 | KNM-ER 3735 | Homo sp. indeterm. |
| 1,8000 | OH7, OH24, OH62, OH65 | Homo habilis |
| 1,8000 | STW 53 | Homo gautengensis |
| 1,8000 | D-211 | Homo georgicus |
| 1,7000 | OH16 | Homo habilis |
| 1,7000 | KNM-ER 406, KNM-ER 732 | Paranthropus boisei |
| 1,6000 | OH13 | Homo habilis |
| 1,6000 | KNM-WT 15000 | Grado erectus |
Parecen posibles las siguientes interpretaciones:
- Australopithecus sediba probablemente desciende de A. africanus, y parece el australopiteco más cercano a Homo.
- Tiene rasgos craneodentales y patrones de locomoción más avanzados que Sterkfontein Stw 53, Olduvai OH 62 y Koobi-Fora KNM-ER 3735, más o menos contemporáneos en el tiempo y asignados a H. habilis, por lo que esta asignación se vuelve incierta.
- Sin embargo, para Kimbel y Rak (2017), MH 1 muestra afinidades inequívocas en su morfología zigomáticomaxilar y supraorbital con los cráneos de Sterkfontein miembro 4.
- Su talón, más primitivo que el de Australopithecus afarensis, da pie a pensar a que forma parte de un linaje diferente al representado por A. afarensis y A. africanus (Thomas C. Prang (2015). El patrón de crecimiento facial es único entre los Hominini tempranos (Lacruz et al, 2015).
- Australopithecus sediba podría estar en el camino evolutivo a Homo ergaster. Esta alternativa es defendida, entre otros, por Chris Stringer.
- Homo habilis constituiría una una rama aislada y sin descendencia del árbol de Homo.
- Dembo et al (2015) defienden que sediba podría clasificarse dentro de Homo.
- Si AL-666-1 no es Homo y Stw 53, OH 62 y KNM-ER 3735 no son habilis, entonces sediba sería anterior y se podría admitir una secuencia filogenética Australopithecus sediba - Homo habilis - Homo erectus. Sediba comparte con erectus algunos rasgos craneales que no se exhiben en habilis.
- En contra de esta hipótesis, la anatomía de los restos candidatos a Homo temprano es diferente a la del sediba.
- El fósil de Homo temprano LD 350-1 es más derivado en su morfología dental y mandibular a pesar de ser anterior en el tiempo en 0,75-0,8 Ma.
- Podría representar un grupo hermano que persistió durante algún tiempo después de la primera aparición de Homo. Ésta parece la hipótesis más probable, defendida entre otros por Donald Johanson.
El dimorfismo sexual en sediba
parece similar al de los humanos modernos.
Ecosistema
Del análisis de los tejidos adheridos a las placas dentales, se deduce que su alimentación incluía cortezas blandas, hojas, frutos y otros productos vegetales de los ambientes boscosos y cerrados, incluso duros, una alimentación generalista y diversa muy diferente a la del resto de los australopitecinos y primeros Homo (Amanda G. Henry y otros, 2012). Sin embargo, de acuerdo con estudios biomecánicos (Ledogar et al, 2016) y la forma del ramus (Ritzman et al, 2016), era incapaz de consumir alimentos duros (pero Daegling et al, 2016). Si Australopithecus sediba no estaba adaptado para consumir alimentos duros, el hecho de hallar rastros de tejidos duros en las placas dentales sugiere la competencia por los recursos con otras especies de Hominini.
Con respecto a la fauna asociada, hasta el momento se han recuperado algunos esqueletos parciales de bóvidos. Las especies sugieren un mosaico de praderas C3 y C4, con fuentes permanentes de agua cercanas, aunque los sedimentos sugieren una vegetación tipo C4 (Holt et al, 2016). Los huesos exhiben fracturas pero no marcas de corte y su origen y proceso de formación parece ser el mismo que el de los fósiles homínidos (Juliet K. Brophy et al, 2013).
Adam van Arsdale: Australopithecus sediba
Enlaces:
Australopithecus sediba: A New Species of Homo-Like Australopith from South Africa. Lee R. Berger et al (2010)
Australopithecus sediba and the earliest origins of the genus Homo. Lee R. Berger, 2012.
Australopithecus sediba and the earliest origins of the genus Homo. Lee R. Berger, 2012.
Colección especial en Science. Abril 2013.
Bibliografía:
Bibliografía:
- Dirks, Paul HGM, et al. "Geological setting and age of Australopithecus sediba from southern Africa." Science 328.5975 (2010): 205-208.
- Pickering, Robyn, et al. "Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo." Science 333.6048 (2011): 1421-1423.
- Henry, Amanda G., et al. "The diet of Australopithecus sediba." Nature (2012).
Te puede interesar:
|
No hay comentarios:
Publicar un comentario