 |
| OH5. Paranthropus boisei. |
Paranthropus boisei, Louis Leakey, 1959.
2,3-1,2 Ma. Los hallazgos en África del Este, anteriores a 2,3 Ma, correspondientes a homínidos robustos, se clasifican dentro de Paranthropus aethiopicus.
El primer Paranthropus boisei apareció en BK, Lower Bed II, Garganta de Olduvai, en 1955: OH3 (Olduvai Hominid 3), con caninos deciduos y una gran corona molar decidua. Pero su taxonomía permaneció incierta hasta que Mary y Louis Leakey descubrieron OH5 en julio de 1959, datado en 1,8 Ma, conocido como Dear boy
(Cascanueces). Se trata de un cráneo de subadulto bien preservado casi completo de 530 cc. En la garganta de Olduvai no se halló ninguna mandíbula, pero en 1964 apareció en la orilla oeste del Lago Natron una mandíbula adulta bien preservada, robusta, con megadontia postcanina (NMT-W64-160, Peninj 1).
En 2013, Manuel Domínguez-Rodrigo et al, comunicaron el hallazgo en 2010 en BK de un conjunto catalogado como OH 80, datado en 1,338±0.024 Ma. Los indicios tafonómicos apuntan a un aprovechamiento por carnívoros. OH 80 es el primer esqueleto parcial hallado en África del Este, clasificado en Paranthropus de forma segura. KNM-ER 1500 (fragmentos poscraneales de Koobi Fora, Kenya), fue asignado a P. boisei por su asociación espacial con un fragmento de mandíbula. Se han relacionado otros restos siguiendo criterios asimismo indirectos.
Otros hallazgos:En 2013, Manuel Domínguez-Rodrigo et al, comunicaron el hallazgo en 2010 en BK de un conjunto catalogado como OH 80, datado en 1,338±0.024 Ma. Los indicios tafonómicos apuntan a un aprovechamiento por carnívoros. OH 80 es el primer esqueleto parcial hallado en África del Este, clasificado en Paranthropus de forma segura. KNM-ER 1500 (fragmentos poscraneales de Koobi Fora, Kenya), fue asignado a P. boisei por su asociación espacial con un fragmento de mandíbula. Se han relacionado otros restos siguiendo criterios asimismo indirectos.
| OH 80-1 a OH 80-7. Los dientes se muestran en una posición anatómica aproximada al esquema de un arco dental. A. Dientes anteriores en vista labial, dientes postcaninos en vista oclusal. B. Vistas labiales de los dientes anteriores. Fotografías de J Trueba y MJ Ortega; composición por JL Heaton. |
- KNM-ER 406. 1969. Turkana (Kenya). 1,7 Ma. Richard Leakey. Se trata de un cráneo completo al que sólo le faltan los dientes. Su capacidad craneal es de 510 cc.
- KNM-ER 732. 1970. Turkana. 1,7 mda. Richard Leakey. El cráneo es similar al OH 5, pero más pequeño con cresta sagital. Capacidad craneana de 500 cc. Presentan un fuerte dimorfismo sexual.
- KGA10-525. 1993. Konso Etiopia. 1,4 mda. A. Amzaye. Se trata de un cráneo, incluyendo una mandíbula. Su capacidad craneana es de 545 cc.
- West Turkana, Kenia. 2,3-1,6 Ma.
- Chiwondo Beds. Malawi. 2,3 Ma.
- Omo, Etiopía. 2,3-1,2 Ma.
- Koobi Fora, Kenia. 2,2-1,4 Ma.
- Olduvai, Tanzania. 1,8-1,2 Ma.
- Peninj, Tanzania. 1,7 Ma.
- Chesowanja, Kenia. 1,5 Ma.
- Konso, Etiopía. 1,4 Ma. En 1997 el equipo de Gen Suwa halló un cráneo asociado a una mandíbula.
 |
| Yacimientos de Paranthropus boisei. Bernard Wood y Paul Constantino (2007). Adaptado de Delson et al (2000). |
Lista de hallazgos.
Omo, Ethiopia- Omo F203-1. 1.40–1.45 Ma. M3 mandibular derecho
- Omo L7A-125. 2.19–2.27 Ma. Mandíbula
- Omo L40-19. 2.36–2.40 Ma. Cúbito derecho
- Omo L74A-21. 2.19–2.27 Ma. Mandíbula derecha
- Omo L427-7. 1.8–2.0 Ma. Mandíbula derecha
- Omo L628-1. 2.27–2.33 Ma. P4 maxilar izquierdo
- Omo L628-2. 2.27–2.33 Ma. M3 mandibular derecho
- Omo L628-3. 2.27–2.33 Ma. M3 mandibular izquierdo
- Omo L628-4. 2.27–2.33 Ma. P4 mandibular izquierdo
- Omo L628-5. 2.27–2.33 Ma. P4 mandibular derecho
- Omo L628-9. 2.27–2.33 Ma. M1 mandibular izquierdo
- Omo L704-2. 2.40–2.52 Ma. P3 deciduo mandibular izquierdo
- Omo L726-11. 2.19–2.27 Ma. P4 maxilar derecho
- Omo L797-1. 2.19–2.27 Ma. Fragmento de P4 mandibular izquierdo
- Omo 47-46. 2.19–2.27 Ma. M2 desgastado mandibular derecho
- Omo 47-1500. 2.10 Ma. M2 mandibular derecho
- Omo 76-37. 2.27–2.33 Ma. Mitad bucal de M3 mandibular izquierdo
- Omo 141-2. 2.27–2.33 Ma. M1 o M2 Maxilar M1 or M2
- Omo 323-896. 2.19–2.27 Ma. Cráneo parcial
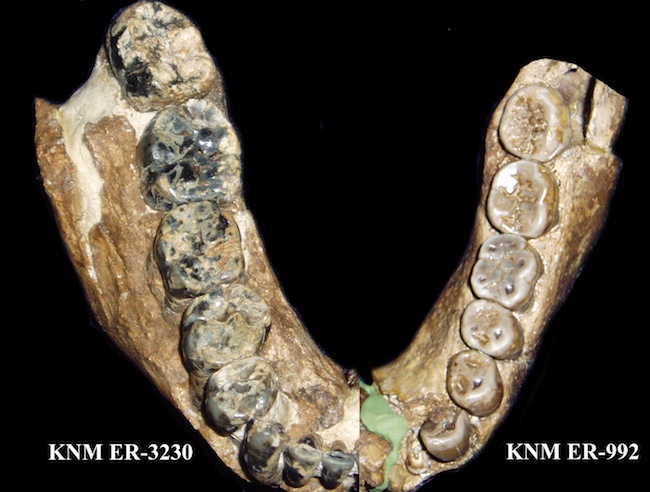 |
| KNM-ER 3230 Paranthropus boisei y KNM-ER 992 Homo ergaster |
- KNM-ER 403. 1.67 Ma. Fragmento mandibular derecho
- KNM-ER 404. 1.49–1.55 Ma. Fragmento mandibular derecho
- KNM-ER 405. 1.87 Ma. Fragmentos palatomaxilares
- KNM-ER 406. 1.56–1.60 Ma. Cráneo sin dientes
- KNM-ER 407. 1.85 Ma. Calvaria
- KNM-ER 725. 1.49–1.55 Ma. Cuerpo mandibular izquierdo
- KNM-ER 726. 1.55–1.65 Ma. Cuerpo mandibular izquierdo
- KNM-ER 727. 1.56–1.60 Ma. Fragmento mandibular derecho
- KNM-ER 728. 1.49–1.55 Ma. Cuerpo mandibular derecho
- KNM-ER 729. 1.49–1.55 Ma. Mandíbula
- KNM-ER 732. 1.56–1.60 Ma. Cráneo parcial
- KNM-ER 733. 1.49–1.55 Ma. Fragmentos de cráneo
- KNM-ER 801. 1.56–1.60 Ma. Fragmento de mandíbula
- KNM-ER 802. 1.56–1.60 Ma. Isolated teeth
- KNM-ER 805. 1.49–1.55 Ma. Fragmento de mandíbula
- KNM-ER 810. 1.77 Ma. Fragmento de mandíbula
- KNM-ER 812. 1.77 Ma. Fragmento de mandíbula
- KNM-ER 814. 1.77 Ma. Fragmentos de bóveda craneal
- KNM-ER 816. 1.77 Ma. Canino maxilar
- KNM-ER 818. 1.55–1.65 Ma. Fragmento de mandíbula
- KNM-ER 1170. 1.56–1.60 Ma. Fragmentos de bóveda craneal
- KNM-ER 1171. 1.56–1.60 Ma. Diente aislado
- KNM-ER 1467. 1.49–1.55 Ma. Diente aislado
- KNM-ER 1468. 1.55–1.65 Ma. Fragmento de mandíbula
- KNM-ER 1469. 1.87–1.90 Ma. Fragmento de mandíbula
- KNM-ER 1477. 1.85–1.90 Ma. Mandíbula sin ramis
- KNM-ER 1479. 1.87 Ma. Diente aislado
- KNM-ER 1500. 1.87–1.90 Ma. Esqueleto parcial
- KNM-ER 1509. 1.56–1.60 Ma. Diente aislado
- KNM-ER 1803. 1.87–1.90 Ma. Fragmento de mandíbula
- KNM-ER 1804. 1.77 Ma. Fragmento de mandíbula
- KNM-ER 1806. 1.85 Ma. Fragmentos de mandíbula
- KNM-ER 1816. 1.56–1.60 Ma. Fragmentos de mandíbula
- KNM-ER 1818. 1.56–1.60 Ma. Diente aislado
- KNM-ER 1819. 1.60–1.87 Ma. Diente aislado
- KNM-ER 1820. 1.67–1.70 Ma. Fragmentos de mandíbula
- KNM-ER 2607. 1.60–1.87 Ma. Diente aislado
- KNM-ER 3229. 1.85 Ma. Fragmento de mandíbula
- KNM-ER 3230. 1.56–1.60 Ma. Mandíbula sin ramis
- KNM-ER 3729. 1.87–1.90 Ma. Fragmento de mandíbula
- KNM-ER 3731. 1.87–1.90 Ma. Fragmentos de mandíbula
- KNM-ER 3737. 1.56–1.60 Ma. Diente aislado
- KNM-ER 3885. 1.77–1.79 Ma. Diente aislado
- KNM-ER 3886. 1.77 Ma. Diente aislado
- KNM-ER 3887. 1.50–1.65 Ma. Diente aislado
- KNM-ER 3889. 1.49 Ma. Fragmento de mandíbula
- KNM-ER 3890. 1.56–1.60 Ma. Diente aislado
- KNM-ER 3952. 1.87 Ma. Diente aislado
- KNM-ER 3954. 1.60–1.87 Ma. Fragmentos de mandíbula
- KNM-ER 5429. 1.56–1.60 Ma. Fragmento de mandíbula
- KNM-ER 5679. 1.56–1.60 Ma. Diente aislado
- KNM-ER 5877. 1.49–1.55 Ma. Fragmento de mandíbula
- KNM-ER 6080. 1.55–1.65 Ma. Diente aislado
- KNM-ER 6082. 1.55–1.65 Ma. Diente aislado
- KNM-ER 6128. 1.77 Ma. Diente aislado
- KNM-ER 13750. 1.87 Ma. Cráneo con endocasto
- KNM-ER 15930. 1.77 Ma. Mandíbula
- KNM-ER 15940. 1.77 Ma. M2 o M3 germinales izquierdos y derechos.
- KNM-ER 15950. 1.49–1.55 Ma. M3 mandibular muy erosionado
- KNM-ER 16841. 1.60–1.87 Ma. Mandíbula sin dientes
- KNM-ER 17760. 1.77 Ma. I2 maxilar izquierdo
- KNM-ER 23000. 1.87 Ma. Cráneo-
- KNM-ER 25520. 1.80–1.90 Ma. Cuerpo mandibular derecho
 |
| KNM-ER 406 |
- KNM-WT 17396. 1.65–1.79 Ma. M3 (o M2) mandibular izquierdo
- KNM-WT 17400. 1.67–1.87 Ma. Craneo anterior y endocasto
- KNM-WT 18600. 1.65–1.79 Ma. P3 maxilar izquierdo
- KNM-WT 37100. 1.65–1.79 Ma. M3 mandibular izquierdo
- KNM-WT 37744. 1.65–1.79 Ma. Canino maxilar izquierdo
- KNM-WT 37747. 1.65–1.79 Ma. M1 maxilar izquierdo
- KNM-WT 37748. 1.65–1.79 Ma. M3 maxilar derecho
- KGA 10-506. 1.41–1.43 Mas. Paladar izquierdo con dentición.
- KGA 10-525. 1.41–1.43 Ma. Cráneo parcial
- KGA 10-565. 1.41–1.43 Ma. M1 maxilar derecho
- KGA 10-570. 1.41–1.43 Ma. Mandíbula juvenil
- KGA 10-900. 1.41–1.43 Ma. Fragmentos molares
- KGA 10-1455. 1.41–1.43 Ma. Parietal izquierdo
- KGA 10-1720. 1.41–1.43 Ma. M3 mandibular izquierdo
- KGA 10-2705. 1.41–1.43 Ma. M2 mandibular derecho
- KGA 10-2741. 1.41–1.43 Ma. Fragmentos molares
- KNM-CH 1. [1.42 Ma. Mitad del rostro y base craneal anterior
- KNM-CH 302. [1.42 Ma. Fragmentos de molar
- KNM-CH 304. [1.42 Ma. 5 fragmentos craneales
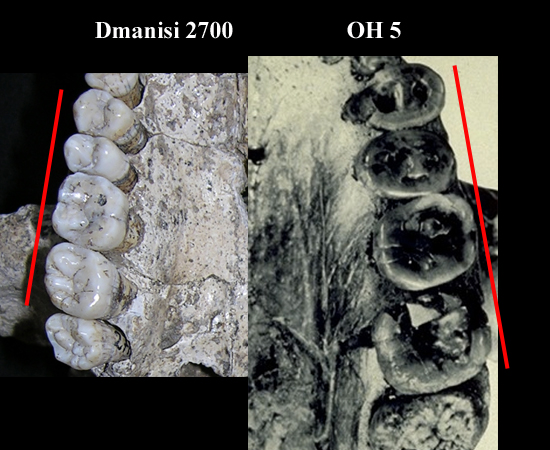 |
| Comparación de OH 5 y Dmanisi 2700 |
- OH 3. 1.40–1.45 Ma. Molar aislado
- OH 5. 1.79–1.85 Ma. Cráneo
- OH 20. 1.66–1.79 Ma. Fragmento proximal del fémur izquierdo
- OH 26. 1.66–1.79 Ma. Molar maxilar sin erupcionar
- OH 30. 1.66–1.79 Ma. Dientes deciduos y permanentes
- OH 36. 1.40–1.45 Ma. Cúbito casi completo
- OH 38. 1.40–1.45 Ma. M2 mandibular and dos incisivos derechos
- OH 46. 1.79–1.85 Ma. Fragmento de corona premolar o molar
- OH 80-1. I2 inferior derecho casi competo.
- OH 80-2. Fragmento de P3 superior derecho unido a un pequeño fragmento de maxilar.
- OH 80-3. Fragmento de P4 superior izquierdo.
- OH 80-4. Fragmento de M1 superior izquierdo.
- OH 80-5. Fragmento de maxilar izquierdo de 5 cm mesiodistalamente, que conserva I1 e I2.
- OH 80-6. I1 superior derecho.
- OH 80-7. I2 superior derecho.
- OH 80-8 y OH 80-9. Raíces extremadamente robustas sin coronas.
- OH 80-10. Porción distal de húmero izquierdo.
- OH 80-11- Fragmento proximal de radio derecho con la mayor parte de su eje.
- OH 80-12. Segmento de la diáfisis de un fémur derecho.
- OH 80-13. Fragmento de eje de una tibia.
- NMT-W64-160. 1.56–1.70 Ma. Mandíbula
- RC 911. 2.3–2.5 Ma. Fragmento de maxilar
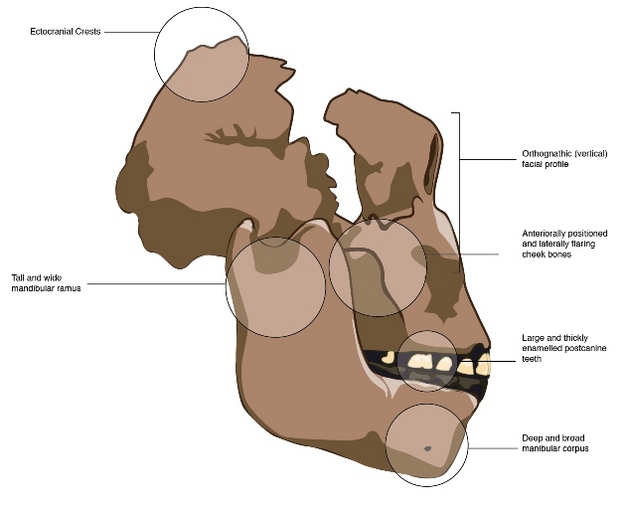 |
| Características de OH5. Nature (2012). |
El problema de los restos postcraneales
La coincidencia en el mismo tiempo y lugar de Paranthropus boisei y Homo temprano dificulta la asignación de los restos postcraneales recuperados. Se asume que:- Un taxón con grandes dientes postcaninos tiene un cuerpo más grande que otro con los postcaninos más pequeños.
- Un especimen que no se parece a los Homo tardíos, pertenece a Paranthropus boisei.
- Cualquier fósil postcraneal hallado cerca de un fósil craneal y derivado del mismo horizonte fosilífero, pertenece al mismo taxón que el especimen craneal.
- Potenciales fósiles procedentes de la Garganta de Olduvai:
- OH 8, pie parcial. Comparte características con TM 1517 (Paranthropus robustus).
- OH 10, falange terminal hallucial.
- OH 20, fémur proximal izquierdo de un adulto, sin la cabeza y puntas de los troncáteres. Hallado en 1959 en HWK, Lower Bed II. No fue asignado a un homínido hasta que Napier reportó Sts 14, SK 82 y SK 97, tres fémures proximales de australopitecinos del sur de África, con los que comparte características.
- OH 35, tibia y fíbula.
- OH 36, cúbito.
- Potenciales fósiles procedentes de Koobi Fora:
- KNM-ER 738, fragmento femoral proximal, 1971.
- KNM-ER 815, fragmento femoral proximal, 1972. Ärea 10.
- KNM-ER 993, fragmento femoral proximal, 1972.
- KNM-ER 1464, astrágalo. Área 6A de Ileret. Características en mosaico, algunas de las cuales recuerdan a Homo. (Anne Su et al, 2013).
- KNM-ER 1465, fragmento femoral proximal. Área 11.
- KNM-ER 1500, esqueleto parcial, 1972. Descubierto en el Miembro Burgi Superior, área 130.
- KNM-ER 1503, fragmento femoral proximal, 1973.
- KNM.ER 1823, terminación proximal del tercer metatarsiano. Área 6A de Ileret.
- KNM-ER 1824, fragmento humeral distal. Área 6A de Ileret.
- KNM-ER 1825, fragmento de atlas. Área 6A de Ileret.
- KNM-ER 5428, astrágalo izquierdo. Área 119.
- ER 739 y ER 1504 fueron atribuidos inicialmente a Australopithecus. ER 1591, a Homo.
- El grado intermedio de aplanamiento anteroposterior está acompañado por un alto grado de convexidad en la superficie posterior (en particular medialmente) que es evidente en la inspección visual (en contraste con la superficie posterior plana observada generalmente en Homo). La morfología de la diáfisis del P. boisei aumenta una lista de características anatómicas del húmero bastante distintivas, incluyendo epicóndilos lateral y medial altamente sobresalientes, un zona conoidea mediolateralmente estrecha, pilares amplios medial y lateralmente y una fosa del olécranon poco profunda (Leakey, 1973; Senut, 1981c; Lague y Jungers, 1996).
- El húmero de OH 80 se caracteriza por un hueso cortical que es considerablemente más grueso que en los H. sapiens modernos (Domínguez-Rodrigo et al, 2013).
- Las diáfisis bien conservados de KNM-ER 739 y ER 1591 muestran pronunciadas marcas musculares.
- Es teóricamente posible que uno o más húmeros de Koobi Fora sin asociación con restos craneodentales correspondan a H. rudolfensis en lugar de a P. boisei (Lague y Jungers, 1996). Sin embargo, los fósiles atribuidos a H. rudolfensis son raros por encima de la tufa KBS (Antón, Potts y Aiello, 2014; Joordens et al, 2013), por lo que la única muestra examinada aquí que podría pertenecer a rudolfensis es KNM-ER 1504. Por otra parte, la gran similitud entre los húmeros de Koobi Fora y la naturaleza distintiva de su morfología articular y diafisaria sugiere que todos ellos pertenecen al mismo género, si no a la misma especie. Dado que KNM-ER 739 es mucho más joven (ca 1,5 Ma) que cualquier H. rudolfensis conocido, y que todos los especímenes Koobi Fora se asemejan a OH 80 en la forma de la diáfisis, lo más probable es que todos estos especímenes representen a P. boisei. Una alternativa (pero menos probable) es que los húmeros de H. rudolfensis sean indistinguibles de los de P. boisei.
- KNM-ER 1504 está asociado con dos fragmentos de fémur: ER 1503 y ER 1505 (Leakey, 1973). El fragmento proximal ER 1503 muestra una morfología primitiva común en los asutralopitecinos y en Orrorin tugenensis: cabeza femoral pequeña, cuello femoral largo y estrecho anteroposteriormente, bajo ángulo cuello-eje, y amplio eje proximal (Richmond y Jungers, 2008; Harmon, 2009; Richmond y Jungers, 2012).
Características
Paranthropus boisei es bien conocido por sus grandes fauces y megadontia. Es todavía más robusto que los parántropos de Sudáfrica. Se reconoce fácilmente y su hipodigma está fuera de controversia. Además, la mayor parte de los fósiles provienen de yacimientos de fácil datación. La evolución de sus variables métricas y no métricas puede trazarse sin dificultad a lo largo de cientos de miles de años (Bernard Wood y Paul Constantino, 2007).
- Autapomorfias
- Sutura parietotemporal superpuesta extensivamente (Rack, 1978). Es una adaptación a los picos de carga de la alimentación (Christine Dzialo et al, 2013)
- Coronas de los premolares mandibulares con talónido particularmente largo y elaborado (Suwa, 1988).
- Corona del primer molar mandibular con cúspides adicionales confinadas al talónido (Wood y Abbott, 1983).
- Simplesiomorfia con expresión distitiva
- Extremada expresión de la megadontia postcanina. El engrosamiento del esmalte es el mayor de todos los hominoideos.
- Combinación distintiva de simplesiomorfias.
- Único homínido que combina una cara amplia, masiva, plana con megadontia postcanina y modesto volumen endocraneal (400-545 cc) (Bilsborough y Wood, 1988). El cerebro del P. boisei no creció a lo largo de los más de 1 Ma que existió la especie (J. Hawks, 2011).
- Otras características:
- Cresta sagital enorme, con un gran polimorfismo, que indica una dieta muy abrasiva.
- Parietales relativamente largos y estrechos, occipital bajo y amplio, huesos de la bóveda craneal muy finos.
- Reducción del tamaño de caninos e incisivos, en línea recta delante del paladar.
- Posición de la espina nasal y aplanamiento de la parte anterior de la cabeza.
- Otros rasgos lo acercan al Australopithecus, como el tamaño del tercer molar superior, más pequeño que el segundo molar.
Se ha sugerido que el grado de variación en el hipodigma mandibular de P. boisei es excesivo para un taxón. Pero, por una parte, los procesos de fosilización podrían alterar el tamaño de los ejemplares y por otra, con independencia del tamaño, todos los ejemplares presentan las mismas características robustas. Para Wood y Lieberman (2001) las regiones sometidas a un alto nivel de esfuerzo, como las masticatorias, tienden a ser más variables.
El grado de variación es consistente con un sustancial dimorfismo corporal combinado con un modesto dimorfismo en los caninos. Con esta información, es difícil hacer inferencias sobre su estructura social.
De los datos producidos a partir de los dientes recuperados, se deduce una ontogenia más parecida a la del humano moderno que a la del chimpancé, si bien el desarrollo de algunos dientes concretos recuerda al chimpancé y el desarrollo de otros dientes no recuerda ni a chimpancés ni a humanos.
El análisis de OH 80 (Domínguez-Rodrigo et al, 2013) proporciona nuevos conocimientos relativos a P. boisei.
El grado de variación es consistente con un sustancial dimorfismo corporal combinado con un modesto dimorfismo en los caninos. Con esta información, es difícil hacer inferencias sobre su estructura social.
De los datos producidos a partir de los dientes recuperados, se deduce una ontogenia más parecida a la del humano moderno que a la del chimpancé, si bien el desarrollo de algunos dientes concretos recuerda al chimpancé y el desarrollo de otros dientes no recuerda ni a chimpancés ni a humanos.
El análisis de OH 80 (Domínguez-Rodrigo et al, 2013) proporciona nuevos conocimientos relativos a P. boisei.
- Se estima una estatura mínima para este individuo de 156 ± 3.91 cm y un peso entre 40-61,7 kg. Esto supone que un indiviuo sustancialmente más grande que el presuntamente femenino KNM-ER 1500, lo que indica un alto grado de dimorfismo sexual.
- Las dimensiones de OH 80-12 quedan dentro del rango de variación de H. erectus, pero son evidentes algunas características distintivas, como un línea espiral mucho más marcada, una tuberosidad glútea más orientada medialmente y un contorno lateral recto de la diáfisis media. Estas diferencias morfológicas probablemente reflejan la variación en la biomecánica de las extremidades inferiores de P. boisei y H. erectus.
- Los análisis morfológicos de OH 80-11 apoyan la afirmación de que el P. boisei tenía las extremidades delanteras relativamente largas y poderosas. OH 80-11 es el hueso del antebrazo homínido más robusto conocido del Plioceno y comienzos del Pleistoceno. Es posible que, perdidas las adaptaciones para la trepa con los primeros Paranthropus, en boisei surgiesen de nuevo como una apomorfia. La morfología radial no es incompatible con la capacidad de agarre (Lague, 2015)-
Izquierda: OH 80-12. Fémur derecho.
Derecha: OH 80-11. Radio derecho.
Ambos fósiles se muestran en vista posterior; con la parte superior hacia arriba.
Barra de escala = 1 cm.
|
Ecología
P. boisei fue uno de los primeros taxones homíndos en explotar paisajes abiertos.
Según Wynn, Cerling y Sponheimer (2013), datos de isótopos de carbono muestran que P. boisei tenían una dieta principalmente de recursos C4 (75%, una dieta mucho menos variada que la de Homo). Estos datos son irreconciliables con la idea de P. boisei consumía alimentos muy similares a los de los simios africanos, frutos secos o frutas duras. Los estudios sobre microdesgaste dental de P. S. Ungar et al (2011) confirman la ausencia de patrones correspondientes al consumo de alimentos duros.
Gabiele A. Macho (2014) considera que pudo tener una dieta similar a la de los babuinos jóvenes que viven en entornos similares y tendrían parecidas necesidades energéticas. Estos babuinos consumen principalmente chufas Cyperus esculentus y eventualmente otros frutos e invertebrados, como gusanos y saltamontes. Esta alimentación habría contenido altas cantidades de minerales, vitaminas y ácidos grasos suficientes para el cerebro homínido. Las chufas, que son ricas en almidones, son altamente abrasivas en un estado sin calentar. El estudio revela también que los dientes de los babuinos tienen marcas de microdesgaste similares a las del boisei. Con el fin de digerir las chufas y permitir que las enzimas de la saliva descompongan los almidones, los parántropos tendrían que masticarlas durante mucho tiempo, presionando el hiperdesarrollo de mandíbulas y dientes.
Han aparecido herramientas líticas en los mismos yacimientos de los que se han recuperado restos asignados a P. boisei y a Homo. Estas herramientas se han asignado a Homo, pero no hay ninguna evidencia de que boisei fuera incapaz de fabricarlas y utilizarlas.
Según Wynn, Cerling y Sponheimer (2013), datos de isótopos de carbono muestran que P. boisei tenían una dieta principalmente de recursos C4 (75%, una dieta mucho menos variada que la de Homo). Estos datos son irreconciliables con la idea de P. boisei consumía alimentos muy similares a los de los simios africanos, frutos secos o frutas duras. Los estudios sobre microdesgaste dental de P. S. Ungar et al (2011) confirman la ausencia de patrones correspondientes al consumo de alimentos duros.
Gabiele A. Macho (2014) considera que pudo tener una dieta similar a la de los babuinos jóvenes que viven en entornos similares y tendrían parecidas necesidades energéticas. Estos babuinos consumen principalmente chufas Cyperus esculentus y eventualmente otros frutos e invertebrados, como gusanos y saltamontes. Esta alimentación habría contenido altas cantidades de minerales, vitaminas y ácidos grasos suficientes para el cerebro homínido. Las chufas, que son ricas en almidones, son altamente abrasivas en un estado sin calentar. El estudio revela también que los dientes de los babuinos tienen marcas de microdesgaste similares a las del boisei. Con el fin de digerir las chufas y permitir que las enzimas de la saliva descompongan los almidones, los parántropos tendrían que masticarlas durante mucho tiempo, presionando el hiperdesarrollo de mandíbulas y dientes.
Han aparecido herramientas líticas en los mismos yacimientos de los que se han recuperado restos asignados a P. boisei y a Homo. Estas herramientas se han asignado a Homo, pero no hay ninguna evidencia de que boisei fuera incapaz de fabricarlas y utilizarlas.
Taxonomía
Leakey optó por
colocarlo en un nuevo género de la familia Australopithecinae (Zinjanthropus boisei)
- Zinj: antiguo nombre de África del Este.
- Charles Boise era el mecenas que pagaba a los Leakey.
Tobias prefirió
incluirlo en el género Australopithecus pero
en el subgénero-especie Paranthropus
boisei. Otros investigadores prefieren utilizar Australopithecus boisei.
Para una justificación de Paranthropus, ver:
El género Paranthropus
.png)
Para una justificación de Paranthropus, ver:
El género Paranthropus
.png)
Adam van Arsdale: Paranthropus boisei
Enlaces:
Paranthropus boisei: Fifty years of evidence and
analysis. Bernard Wood and Paul Constantino. 2007.
Diet of Paranthropus boisei in the early Pleistocene of East Africa. Thure E. Cerling et al (2011)
Bibliografía:
Diet of Paranthropus boisei in the early Pleistocene of East Africa. Thure E. Cerling et al (2011)
Bibliografía:
- Leakey, Louis SB. "A new fossil skull from Olduvai." Nature 184.4685 (1959): 491-493.
- Fleischer, R. L., et al. "Fission-Track Dating of Bed I, Olduvai Gorge." Science 148.3666 (1965): 72-74.
- Leakey, Mary Douglas. Olduvai gorge. Eds. Phillip V. Tobias, and L. S. B. Leakey. Vol. 2. Cambridge University Press, 1967.
- Blumenschine, Robert J., Ian G. Stanistreet, and Fidelis T. Masao. "Olduvai Gorge and the Olduvai Landscape Paleoanthropology Project." Journal of Human Evolution 63.2 (2012): 247-250.
Te puede interesar:
|
No hay comentarios:
Publicar un comentario