Las excavaciones en la cámara central de Denisova, en los Montes Altai, han revelado ocupaciones del Pleistoceno entre 125-30 ka años, con restos líticos musterienses y levalloisienses. Una datación de los huesos de fauna de la parte inferior de la capa 11, realizado en 2015 por Katherina Douka et al ha proporcionado unas fechas de 48-50 ka.
El fragmento de falange Denisova 3 fue examinado por Johannes Krause, Svante Päabo y colegas del Proyecto Genoma Neandertal en la Instituto Max Planck de Antropología Evolutiva. El ADNmt es diferente al de los seres humanos anatómicamente modernos, y al de los neandertales. A falta de otra denominación, se ha convenido tácitamente que perteneció a un "denisovano".
El ADN de varios individuos cuyos restos fueron recuperados en la cueva Denisova permite conocer que la cueva fue habitada por denisovanos entre hace 287-55 ka y por neandertales entre hace 193-97 ka (Jacobs et al, 2019). Los estudios paleogenéticos han revelado que el ADN denisovano se puede encontrar en algunas poblaciones vivas, particularmente en Melanesia y el norte de Australia y han calculado las fechas de divergencia con los neandertales y los humanos modernos y de diversos eventos de hibridación. Denisovanos y neandertales divergieron de un antepasado común hace 390 ka. Entre ambos grupos hubo episodios de hibridación. Asimismo, los denisovanos se mezclaron con los antepasados de los actuales humanos de Oceanía y Asia. Por otra parte, los denisovanos recibieron el aporte genético de uno Homo arcaico que divergió hace más de 1 Ma de los antepasados de los humanos actuales.
Se han recuperado muy pocos restos de Homo, entre ellos (dataciones de las capas estratigráficas de Jacobs et al, 2019),
Denisova 2. Segundo molar inferior deciduo muy desgastado recuperado en 1984 en la capa 22.1 de la Galería Principal, datada en más de 287 ka. Según el análisis genético (ADNmt y ADN nuclear), perteneció a una niña denisovana que vivió entre hace 139-184 ka (Slon et al, 2017). La morfología indica una edad de 10-12 años.
Denisova 3. En 2008, un equipo dirigido por los arqueólogos Michael Shunkov y Anatoli Derevianko de la Academia Rusa de Ciencias de Novosibirsk encontró en la capa 11.2 de la cámara Este, una banda gruesa de piedra arenisca, una fragmento distal de falange de un infantil. La capa se ha datado en más de 55 ka, mientras que la datación por medios genéticos es de hace ca. 85 ka.
En 2010, el equipo de Max Planck secuenció el 70% del genoma nuclear. El genoma quedó completado en 2012.
Según los análisis genéticos, la falange perteneció a una joven denisovana de pelo y ojos castaños y tez oscura. La población era reducida y muy endogámica.
El genoma incluye trazas de un homínido desconocido que divergió hace 4-1 Ma del ancestro de denisovanos, neandertales y HAM.
Según estos estudios, completados por el equipo de David Reich (2011), el ADN de los denisovianos comparte ciertas mutaciones con el de los melanesios de Papúa Nueva Guinea y la isla de Bougainville, los aborígenes australianos, los nativos de Fiji, los indonesios del este, y los Mamanwa (Filipinas) que no se encuentran en los neandertales u otras poblaciones modernas. Tampoco se encuentra en los asiáticos del este del continente, los indonesios occidentales, los Jehai (Malasia), y los Onge (Islas Andamán). Estos resultados sugieren que los denisovanos vivieron dentro de una extraordinariamente amplia gama geográfica y ecológica, desde Siberia hasta el Asia tropical y que los ancestros de los asiáticos orientales no estaban en el sudeste de Asia, cuando se produjo el flujo de genes denisovanos.
Denisova 4. Créditos: Bence Viola.
Denisova 4. Molar de macho adulto hallado en la capa 11.1 de la cámara Sur (menos de 47 ka). Un análisis del ADNmt confirmó su adscripcion denisovana (Viviane Slon et al, 2015). El ADN nuclear fue secuenciado por Sawyer et al, 2015.
Denisova 5. Falange del pie hallada en la capa 11.4 de la cámara Este (más de 105 ka) que perteneció a un neandertal, en base al análisis genético. La datación por medios genéticos es de hace ca 123 ka.
Denisova 8. Probablemente un M3 superior de macho adulto, hallado en 2010 entre las capas 11.4 y 12.1 (120-105 ka). Es similar a Denisova 4 en su gran tamaño y en el mosaico de rasgos de HAM y neandertales que presenta. El ADN nuclear y mitocondrial fue secuenciado por Sawyer et al, 2015 y corresponde a un denisovano.
Denisova 9. Capa 12.2 de la cámara Este. 141-129 ka. Neandertal.
Denisova 11. Fragmento de hueso largo de una mujer de al menos 13 años de edad, hallado en la capa 12.3 de la cámara Este, datado en menos de 141 ka. El ADN-mt corresponde a un neandertal (Brown et al, 2016), pero el ADN nuclear ha revelado que el padre del individuo era denisovano, con algún ancestro neandertal. La madre procedía de un población neandertal diferente a la del ancestros del padre, más cercana a neandertales europeos posteriores que a los neandertales más antiguos procedentes de la misma cueva. (Slon et al, 2018). La datación genética es de hace más de 90 ka.
Denisova 13. Se ha anunciado el hallazgo en 2016 de un fragmento de parietal con ADN-mt denisovano en la capa 22, datada en hace 280-250 ka.
En la campaña de 2020, se recuperaron un fragmento de hueso en la capa 13 y otro en la 14 (hace 120-130 ka). En la capa 22, un molar en la parte superior (170-190 ka) y un diente deciduo en la parte inferior (250 ka).
Estratigrafía y dataciones de Denisova. Los conjuntos de artefactos están indicados a la derecha de cada secuencia. Las condiciones climáticas se han inferido de los registros de polen y fauna. Dataciones estimadas según un modelo bayesiano con el 95,4% de probabilidad. Jacobs et al, 2019.
Slon et al (2017) han conseguido detectar ADNmt denisovano y neandertal en las capas más profundas de la Cueva Denisova, en concreto de neandertales en la capa 14 y de denisovanos en la capa 15.
Comparación de Denisova 4 y 8 con M3 superior de neandertal y HAM. Sawyer et al, 2015.
Denisova 8. Sawyer et al, 2015.
En la galería este, aparecieron varios objetos ornamentales, entre ellos un anillo de mármol, un brazalete de piedra verde, una diadema de marfil de mamut lanudo y dientes perforados, agujas y colmillos de mamut trabajados, de autoría dudosa. En 2018 restos de ocre y en 2019 la capa 11 (40-45 ka, con restos denisovanos) proporcionó un fragmento de figurilla de león en marfil alóctono y posiblemente pintada con ocre rojo, que es candidata a la representación zoomorfa más antigua.
Cronología de fosiles asiáticos de Homo arcaico. Kaifu, 2017.
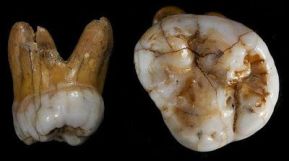
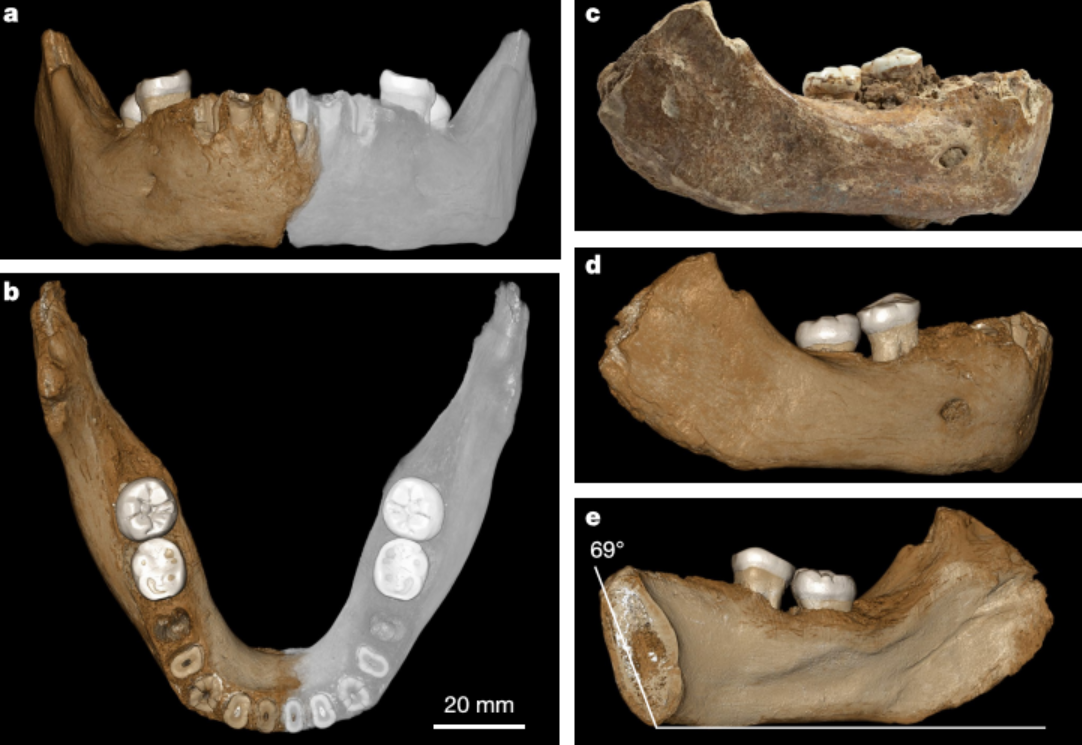
En 2019, Chen et al presentaron el hallazgo de una mandíbula, identificada como denisovana por análisis de proteínas antiguas, en la cueva de Karst Baishiya, Xiahe, Gansu, meseta tibetana, China, datada en hace al menos 160 ka mediante series U de una matriz de carbonato adherida. Por consiguiente, los denisovanos se adaptaron con éxito a ambientes hipóxicos de gran altitud mucho antes de la llegada a la región del HAM.
La mandíbula presenta un segundo molar inferior de tres raíces. Esta característica es rara en el sapiens no asiático, pero en poblaciones de origen asiático puede superar el 40% de ocurrencia. Existe pues la posibilidad de que la mayor frecuencia de la presencia de este rasgo en los asiáticos se deba a una introgresión denisovana (Bailey et al, 2019).
Según María Martinón-Torres et al (2017), los dientes denisovanos se caracterizan por su tamaño notablemente grande y sus rasgos morfológicos primitivos (Krause et al, 2010; Sawyer et al, 2015). A pesar de la estrecha relación filogenética entre los denisovanos y los neandertales, los molares denisovanos no muestran ningún rasgo neandertal típico. Estos dientes caen dentro del rango de variación de los fósiles del Pleistoceno Temprano y Medio del este de Asia en las cúspides mesiales bucolingualmente expandidas, la esquina distovestibular decreciente y las raíces divergentes y gruesas. Esta morfología podría interpretarse como una supervivencia tardía de un patrón típicamente asiático o como resultado de un mestizaje genético con una especie desconocida (tal vez un H. erectus superviviente ). Además, el tamaño dental está dentro del rango de los homininos del Plioceno y del Pleistoceno Temprano, lo que podría evidenciar un caso de heterosis, una consecuencia fenotípica de la hibridación genética (Ackermann, 2010; Ackermann, Rogers y Cheverud, 2006).
Otras muestras del registro fósil asiático podrían pertenecer a denisovanos, en particular Xujiayao, Penghu 1, Maba y Xuchang (María Martinón-Torres et al, 2017).
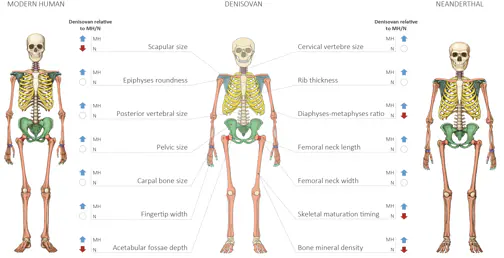
Comparación del esqueleto de HAM, denisovano y neandertal. Gokhman et al (2019).
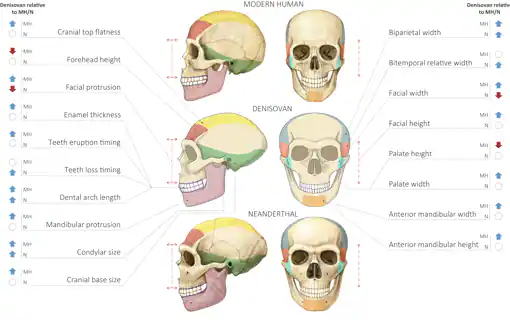
Comparación de los cráneos de HAM, denisovano y neandertal. Gokhman et al (2019).
En ausencia de fósiles que permitan especular sobre la morfología denisovana, Gokhman et al (2019) han utilizado los patrones de metilación del ADN. De acuerdo con sus resultados, los denisovanos compartían con los neandertales una cara alargada y una pelvis ancha. Como rasgos distintivos, los autores señalan un aumento del arco dental y la expansión craneal lateral. El cráneo de Xuchang, de difícil clasificación, muestra estos rasgos.
Objetos ornamentales hallados en la Cueva Denisova. Institute of Archaeology and Ethnography SB RAS.
Ubicación de la Sierra de Atapuerca. G. Cuenca-Bescós et al., 2004
Ubicación
La Sierra de Atapuerca (Burgos, España) es un karst que contiene medio centenar de cavidades completamente colmatadas. Estas cuevas se originan a partir de conductos subhorizontales con paleodrenajes en sentido SN, localizándose la zona de descarga en la cabecera del río Pico. Los conductos están organizados en tres niveles principales que aparecen colgados entre 90 y 60 m sobre el actual cauce del río Arlanzón, coincidiendo con los niveles de base generados por sus terrazas fluviales T2, T3 y T5. La incisión fluvial liberó de las aguas los conductos superiores mientras se excavaban los niveles inferiores del karst. Las cuevas que iban quedando accesibles fueron utilizadas por la fauna y los homininos, conservando un registro arqueo-paleontológico de más de 1,2 Ma. Las intervenciones más importantes han sido las siguientes:
Cuevas:
Nivel superior del karst.
Galería del Sílex.
Cueva Mayor.
El Portalón. Entrada a Cueva Mayor.
Salón del Coro.
Galería de las Estatuas.
Nivel intermedio del karst.
Sala de los Cíclopes.
Cueva del Silo. Conectada por un estrecho pasaje con Cueva Mayor.
Sima de los Huesos, al final de la Galería del Silo.
Galería Baja. Remata en el relleno de Trinchera Elefante.
Trinchera del Ferrocarril. Este conjunto vio la luz cuando la sierra fue atravesada por un tren minero a finales del XIX.
Sima del Elefante o Trinchera Elefante.
Galería o Trinchera Galería, que incluye la Covacha de los Zarpazos.
La Paredeja. Es una cueva colmatada de sedimentos que una cantera dejó al descubierto. En los nivles superiores, se ha recuperado un rico conjunto de herramientas de sílex y cuarcita realizadas por grupos de neandertales que habitaron el extremo sur de la sierra de Atapuerca, aprovechando los recursos líticos que este ambiente les proporcionaba. La Paredeja es un lugar estratégico desde el que se divisa el valle del río Arlanzón y que estuvo ocupado por estos grupos de cazadores y recolectores.
La Sierra de Atapuerca es zona de paso hacia diferentes ecosistemas, con fáciles accesos desde las vegas de los ríos Pico, que nace en la propia sierra, Vena, que la bordea por el NE, y Arlanzón, Que discurre al SW, por Ibeas de Juarros, a 930 m, por lo que la sierra apenas se eleva 150 m sobre su cauce.
Constituye la estribación más noroccidental de la Cordillera Ibérica (Sierra de la Demanda), estando separada del borde meridional de la Cordillera Cantábrica (Sierra de Ubierna) por un corredor tectónico (Estrecho de Burgos) que enlaza las depresiones terciarias de la Cuenca del Duero y de La Bureba, ésta ya perteneciente a la Cuenca del Ebro.
Las paredes de Cueva Mayor conservan inscripciones que se remontan al siglo XV, al menos. Dos manuscritos relatan las visitas efectuadas a la cavidad en el XVI.
En 1795, Manuel Francisco de Paula relata que un grupo de vecinos de Rubena realiza la exploración sistemática de Cueva Mayor y Cueva del Silo, descendiendo a todas sus simas, entre ellas la Sima de las Huesos, extrayendo los primeros restos fósiles de los que tenemos constancia escrita.
En 1863, Felipe de Ariño solicitó la cesión de la propiedad de Atapuerca por 60 años, con la finalidad de hacer investigaciones en su interior, así como de realizar un plano de la misma. Felipe de Ariño y Ramón Inclán publicaron el hallazgo de restos humanos en Cueva Ciega.
En 1864, Casiano de Prado incluye en su catálogo de cuevas españolas las cavidades de Atapuerca y Ojo Guareña.
En 1868 Aparece publicada la Descripción con planos de la Cueva Llamada de Atapuerca (Pedro Sampayo y Mariano Zuaznávar, 1868), que incluye una guía descriptiva y analiza los aspectos morfológicos y geológicos, así Como algunos de los arqueológicos y paleontológicos.
En 1890 se otorga la concesión minera de la Cueva de Atapuerca a Ramón Inclán. En la solicitud, se incluía una copia de los planos de Sampayo y Zuaznávar a la que se habían añadido los perfiles de El Silo (Sima de los Huesos) y La Galería de las Estatuas.
Entre 1896-1901 La Sierra Company Limited construyó una línea férrea de vía estrecha desde Monterrubio de la Demanda a Villafría, a las puertas de Burgos. La idea era transportar el carbón y el mineral de hierro desde la Sierra de la Demanda hasta el enlace con la línea Burgos-Bilbao. Hacia 1910, la línea férrea dejó de funcionar. Las obras dejaron al descubierto varios yacimientos, al atravesar las estribaciones de la Sierra de Atapuerca.
En 1910 Jesús Carballo describió restos cerámicos, óseos, faunísticos, algunos de especies ya extinguidas, y muestras de arte rupestre de la Cueva Mayor.
En 1912 Henri Breuil y Hugo Obermaier profundizaron en el estudio del arte rupestre de la Cueva Mayor, desmintiendo alguna de las figuras publicadas por Carballo y describiendo nuevos motivos en diferentes galerías, así como en la Cueva del Silo (Breuil y Obermaier, 1913).
En 1926, el geólogo José Royo y Gómez cita por vez primera (Royo, 1926) los rellenos pleistocenos de la Trinchera. También describe Cueva Mayor y Cueva del Silo.
Entre 1930 y 1936, aproximadamente, pueden datarse dos croquis de Cueva Mayor y Cueva del Silo realizados por José Ruiz Goyo. Gracias a ellos, podemos vislumbrar los importantes cambios provocados a mediados del siglo XX por la cantera localizada junto a su entrada (Ortega, 2009).
Una cantera, en la década de 1950, tropezó en su avance con una gran cavidad, casi colmatada de sedimentos, la Cueva del Fantasma. Se trabaja en ella desde 2015.
La exploración y estudio de cavidades en la provincia de Burgos se reanudó en 1951, con la creación del Grupo Espeleológico Edelweiss. En 1954 el grupo comienza sus trabajos en la Sierra de Atapuerca. En 1962, bajo la dirección de José Luis Uribarri, se descubren los primeros restos de fauna fósil en el yacimiento de la Trinchera del Ferrocarril de Atapuerca (Osaba, 1963).
En compañía del director del Museo Arqueológico Provincial de Burgos, Basilio Osaba, y de diversos periodistas y fotógrafos, el 15 de abril de 1963, se realizaron diversas catas en las que apareció un bifaz achelense de cuarcita verdoso que sirvió para contextualizar el yacimiento en el Paleolítico Inferior (Osaba, 1965). Posteriormente indicaron la existencia del yacimiento, tanto a Miquel Crusafont como a Francisco Jordá.
SRB, SRM, SRA: Pozos de sondeo en Rampa Baja, Rampa Media y Rampa Alta.
Descripción
La Trinchera del Ferrocarril es una zanja artificial excavada durante las obras de trazado de la vía férrea. Forma un arco de 500 m de longitud que discurre en dirección NS por la parte meridional de la Sierra. No supera los 20 m de altura y en ella se localizan cavidades seccionadas rellenas de sedimentos de distintos orígenes. Tres de ellas constituyen yacimientos arqueológicos. De norte a sur:
21 unidades sedimentarias agrupadas en tres fases:
Fase III: TE21-TE20.
Colmatación el el Pleistoceno Superior. Estéril.
Fase II: TE19-TE15. Se han recuperado 41 restos líticos, 36 de ellos en TE19, de Modo 2.
TE19-TE18 (Trinchera Elefante Upper Red Unit, TE-URU) se ha datado a partir de datos biocronológicos en 350-250 ka (MIS 9-8), El análisis por series de uranio de una costra estalacmítica recuperada en el techo de TE18 ha aportado dataciones de 307 y 255 ka. Esto sugiere que TE19 tiene una cronología posterior a 255 ka. Contienen abundantes restos de fauna. TE19, con un amplio conjunto de grandes mamíferos asociados con útiles Modo 2.
TE17-TE15. Entrada masiva de depósitos miocenos del cercano Valle de la Propiedad.
TE 17 se ha datado por luminiscencia en hace 781 ka.
En la base de de la TE17 se da un cambio de polaridad. Los sedimentos por debajo corresponden al subcrón Matuyama (> 780 ka).
TE16 se ha datado por lumiscencia en hace 864-804 ka.
Fase I: TE14- TE7. Unidad Roja Inferior. Lower Red Unit (TE-LRU). Presenta una asociación faunística de micromamíferos logomorfos, aves y castores junto con animales de talla mediana y grande, que indican una edad pre-Jaramillo.
El Nivel TE9c se ha datado por análisis de núcleos cosmogénicos en 1,22 Ma.
Se han recuperado una mandíbula, una falange y un fragmento de húmero asignados incialmente un Homo antecessor y actualmente a Homo sp., asociados a industria lítica Modo 1.
El Nivel TE9d se ha datado en ca 1,3 Ma.
Se han recuperado huesos de fauna con marcas de corte y un fragmento de lasca de sílex.
Se han Recuperado 86 artefactos líticos, en su mayoría de sílex (33 corresponden a TE9c). La materia prima provenía de las proximidades. Una lasca de sílex procede de TE8, evidenciando la actividad antrópica más antigua hallada en Atapuerca. El conjunto corresponde al Modo 1.
Más de diez mil restos de fauna (Faunal Unit 1). Algunos de ellos, en su mayoría de ungulados en las unidades TE-13, TE-12 y TE-9, presentan marcas de corte y fracturación. También se ha identificado actividad antrópica en animales pequeños, como aves, tortugas y conejos. El acceso fue primario. Los restos apuntan a ocupaciones de baja intensidad en zonas próximas a la entrada de la cavidad.
En la campaña de 2019 las dos áreas en las que se venía excavando se han unido en la superficie del nivel 7, siendo este el único nivel que han excavado hasta el momento en toda la superficie del yacimiento, con una cronología alrededor de 1,4 Ma, con la recuperación de los restos de un individuo inmaduro y las extremidades superiores de un adulto en conexión anatómica, así como restos de fauna que confirman un ambiente templado y húmedo.
Estratigrafía y dataciones de Gran Dolina. José María Bermúdez de Castro et al, 2013.
Secuencia sedimentaria de 18 m de altura, más otros 8 m sondeados en su base.
Hace ca 1 Ma Gran Dolina sufrió un derrumbe de su techo, convirtiéndose en una trampa de más de 15 m de profundidad.
Unidades sedimentarias:
TD11
Colmatado del depósito hace 240 ka con formación de terra rosa en las grietas del techo hace 55 ka.
Arqueológicamente estéril.
TD10
La mayor acumulación de restos arqueológicos de Atapuerca. 97.000 restos de fauna y 37.000 de industria lítica. Estos materiales son el producto de una sucesión de fases de ocupación muy ricas, con características muy diversas: desde verdaderos campamentos base diversificados a campamentos especializados de cazadores de bisontes, pasando por fases donde las ocupaciones humanas son de un carácter más esporádico. Diversidad y estandarización de las secuencias de explotación. Son predominantes los métodos centrípetos de extracción de lascas. Los sedimentos del nivel TD10 se fueron depositando lentamente a lo largo de 200 ka, un período en el que la cueva de la Gran Dolina estuvo habitada por humanos durante largos períodos de tiempo.
Datado por ESR en hace 450-350 ka.
TD10.1
Los hallazgos líticos en la base podrían representar la evolución local del Modo 2 al Modo 3.
Amplio espectro de presas.
TD10.2
Caza especializada de Bison sp. Cazadero utilizado durante varios eventos estacionales de caza comunal en los que rebaños completos de bisontes fueron sacrificados para ser explotados intensamente. El repetido uso estacional de un punto en el territorio para el desarrollo de tareas específicas muestra ciertas similitudes con el patrón logístico de gestión de los recursos y nos informa sobre la emergencia de habilidades cognitivas, tecnológicas y sociales.
TD10.3
El tramo medio se ha datado en unos 0,45 Ma de antigüedad, documentando una menor concentración de restos con evidencias de actividad humana que en las capas más altas. Han aparecido pocos instrumentos líticos, y los huesos con marcas de procesado por parte de Homo son escasos. Destaca la alta representación de grandes carnívoros.
TD10.4
La excavación de la parte inferior de TD10 proporcionó una gran acumulación de industria lítica, en la que destaca una proporción inusual de bifaces. Datado en hace algo más de hace 400 ka.
TD9
Nivel de limos y guano formado en condiciones de cueva cerrada.
Datado por termoluniniscencia en hace 480 ka.
Algunas piezas líticas.
TD9/8.
Sin evidencias de cultura material.
TD8.
Sellado por espeleotema fechado por series de uranio entre hace 450-400 ka. Edad ponderada de hace 602 ka.
Serie de coladas de cantos que colmatan parte de la sección.
Acumulación faunística con predominio destacado de gamos (Dama vallonnetensis) y presencia ocasional de carnívoros. Marcas de mordedura de carnívoros. Varios tipos de ocupaciones incluyendo guarida de hiénidos.
No hay evidencias de cultura material.
TD7.
En el techo de la unidad, se encuentra el límite paleomagnético Matuyama-Brunhes.
Depósito fluvial.
Restos de Stephanorhinus etruscus y Praeovibos en conexión anatómica, lo que sugiere una posible reactivación de la trampa natural.
TD6-2.
Dataciones por ESR proporcionaron una edad entre hace 0,85-0,77 Ma para el límite superior de TD6. Análisis de lumniscencia, proporcionaron una edad de TD6 de hace 846 ± 57 ka.
Ha proporcionado alrededor de 160 huesos de unos 11 individuos que han permitido la definición de Homo antecessor, así como más de 300 artefactos líticos de sílex, cuarcita, arenisca, caliza y cuarzo, de tecnología Modo 1 y fósiles de mamíferos. Los depósitos sugieren un entorno con un arroyo que cruza la cueva por el medio de la cavidad, adaptándose al paleo-relieve, inundando la parte sureste de la cueva. Con el tiempo, las entradas de flujos de escombros de una entrada sub-vertical al noroeste interrupieron la corriente.
Se distinguen tres grupos de fósiles de Homo:
Un grupo de fósiles dispersos en la facies D1 de la capa TD6.2.2 Torreón, en el noroeste y en la facies F de las capas TD6.2.2 y TD6.2.3.
TD6.2.2 está formada por escombros alóctonos que se introdujeron en la cueva por una entrada sub-vertical. Los fósiles de Homo se hallaron muy juntos en la parte noroeste.
TD6.2.2 y TD6.2.3 son depósitos sedimentarios procedentes de llanuras de inundación y escombros. Los fósiles de Homo aparecieron dispersos.
Tanto los fósiles de Homo como los otros restos arqueológicos, fueron arrastrados por los procesos de flujo de escombros desde el exterior.
Fósiles en la facies A, capas TD6.2.2, TD6.2.1 y TD6.1.2.
La facies A se formó gracias debido a un flujo de canal que arrastró gravas de piedra caliza al interior de la cueva. Los fósiles de Homo se incluyen en las gravas, y su tamaño está dentro del intervalo de la grava.
Fósiles en la facies F. Este grupo constituye la acumulación principal, en la capa TD6.2.
La facies F es el resultado de dos procesos geológicos.
Sedimentos procedentes de una llanura de inundación que entraron en la cueva por la entrada principal de la cueva, al noroeste.
El entorno de inundación no tuvo suficiente energía para arrastrar clastos calizos o restos fósiles.
Flujo de escombros que se introdujeron en la cueva por una entrada secundaria ubicada al sur y que arrastró clastos y fósiles.
Por consiguiente, la actividad de los Homo se habría desarrollado fuera de la cueva, pero muy cerca de ella: la excelente conservación tafonómica de los huesos indica un tiempo muy corto a la exposición externa, un transporte rápido y en distancias muy cortas. Una consecuencia de esta hipótesis es que la datación de los correspondientes fósiles podría ser más antigua de la del nivel, que se ha datado en 960 ka.
Los restos de fauna presentan marcas de corte y fracturas intencionales, incluyendo huesos del propio antecessor, lo que indica canibalismo.
La industria lítica utiliza todas las materias primas aptas del entorno, con todas las fases representadas de la cadena de producción. Estrategias de explotación variadas. Aparece el retoque por vez primera en Atapuerca.
TD6-3 y TD5.
Restos faunísticos. Sobre todo hiénidos que ocuparon la unidad como cubil, alternado con osos.
Restos de ocupación de Hominini, Pero en menor medida.
TD4-3.
Dataciones por ESR proporcionaron una edad 0.91 ± 0.25 Ma para el límnite inferior de TD4.
Restos faunísticos, en particular, de Ursus dolinensis. La cueva era su lugar de hibernación y algunos ejemplares cayero a la sima. También restos de ungulados, algunos con marcas de corte, lo que indica que los humanos realizaban accesos primarios sobre los animales que se precipitaban a la sima.
Pequeña colección de artefactos de cuarcita con talla sencilla.
TD2-1.
Ambiente de cueva cerrada durante una fase de polaridad inversa, con un pequeño intervalo normal relacionado con el subcron Jaramillo o Cobb Mountain.
Modo 2 asociado a abundantes restos faunísticos. 7 tipos de materia prima obtenidas en fuentes situadas a menos de 5 km. Cadenas operativas muy fragmentadas. Gran parte de la talla es alóctona. La talla autóctona es expeditiva. Instrumentos utilizados principalmente para carnicería, pero también para trabajo sobre piel y vegetales.
La mayoría de los restos faunísticos pertenecen a herbívoros, con marcas de carnívoros y en menor medida de humanos. Dos restos humanos en GIII-GII.
Se trata de una cueva a la que accedieron los seres humanos y los carnívoros, por una entrada 2 m al oeste de la Covacha de los Zarpazos, con el fin de obtener la biomasa animal de los herbívoros que habían caído en la cueva a través de un eje natural (TN). El registro arqueológico es incompleto y fragmentado, ya que es producto de un dinámica muy cambiante.
Fragmento de mandíbula y fragmento craneal asignados a Homo heidelbergensis.
Dividido en tres sectores.
Tramo central (TG).
Covacha de los Zarpazos (TZ) al norte.
Trinchera norte (TN) al sur. Conducto vertical abierto al exterior.
Cinco unidades litoestratigráficas y un paleosuelo.
GV
Colmatado de la cueva.
GIV
El espeleotema que sella la unidad y la secuencia en el sector TG se ha datado en 118 y 200 ka por series de uranio y ESR rerspectivamente.
Más de un centenar de restos entre líticos y faunísticos.
GIII
Utilización humana esporádica para aprovechamiento de recursos cárnicos. La torca de TN actuó como una trampa natural para los hervíboros, presentado sus carcasas un acceso diferencial entre los cánidos y los humanos.
GIIIa, datada en 0,30-0,22 Ma, con abundantes restos de fauna, principalmente de caballos y ciervos pero también de carnívoros, e industria achelense. En la parte inferior, se han recuperado restos de fauna e industria lítica Modo 2 principalmente en sílex. También se han recuperado cantos de cuarcita que fueron utilizados como percutores o machacadores de huesos y un núcleo de sílex neógeno de grandes dimensiones. Esta industria Achelense se caracteriza por la presencia de bifaces y pequeñas lascas retocadas (puntas y raederas).
GII.
Datada en 500 ka.
Industria lítica Modo 2, la más antigua de Atapuerca.
GI
Arqueológicamente estéril.
En la base, se ha detectado la transición Matuyama-Brunhes.
20 m por encima de la Trinchera del Ferrocarril se encuentra la Cueva del Fantasma, el yacimiento más amplio de la Sierra, con unos 300 m2. Algunas herramientas recuperadas pertenecen al Musteriense.
A menos de 1 km de la Trinchera del Ferrocarril está situado el complejo Cueva Mayor (Cueva de Atapuerca)-Cueva del Silo (Cueva del Valhondo), que con 3.700 m topografiados, es una de las mayores cavidades conocidas en la Cuenca del Duero.
La cavidad se estructura en tres niveles: Cueva Mayor da acceso a los dos superiores, mientras que Cueva del Silo lo hace al nivel más bajo.
La entrada de Cueva Mayor se abre en el fondo de una fractura, dando acceso a un cómodo Portalón.
Desde Cueva Mayor se accede a una amplia galería de dirección SSE-NNW, cuyo primer tramo es conocido como Salón del Coro. Tras dejar al W el acceso al nivel intermedio, se abre la Galería de las Estatuas.
El nivel intermedio es de dimensiones sensiblemente menores. En dirección NNW recibe el nombre de Galería Baja y en en dirección SSE el de Galería del Silo. En el tramo central de la Galería Baja se observa la existencia de una antigua entrada colmatada. La Galería termina en el relleno de la Sima del Elefante, en la Trinchera, otra antigua entrada colmatada.
Tras superar una estrechez, la Galería del Silo permite acceder a la Sala de los Cíclopes, en cuyo extremo SW se encuentra el Tubo de los Vientos, punto de enlace con la Cueva del Silo. En el extremo SE de la sala, próxima a otra antigua entrada colmatada, se abre la Sima de los Huesos, conocida anteriormente como El Silo, a 500 m de la entrada de Cueva Mayor. Se trata de una sima vertical de 13 m de profundidad: una rampa inclinada de 10 m de longitud que salva un desnivel de 5 m para llegar a una pequeña cámara y una cámara de 8 m de longitud y 3 de anchura.
Por el otro extremo del Portalón se accede a la Galería del Sílex, el conducto más amplio de toda la cavidad.
Estratigrafía, plano y sección de la Sima de los Huesos. Sala et al, 2016.
Sima de los Huesos (Aranburu, Arsuaga y Sala, 2015). Se encuentra en la base de un pozo de unos 13 m de profundidad, a unos 500 m de la actual entrada de Cueva Mayor.
Unidad aloestratigráfica AU-5
LU-12. Bloques de piedra caliza desprendidos del techo de la cavidad.
LU-11. Similar en composición a LU-7 y LU-9. Rico en guano de murciélago. Originado por caída de sedimentos desde las chimeneas. Datación similar a LU-7.
LU-10. Colada extensa. Indica un hiato en la deposición de sedimentos. Formada por el agua que gotea del techo cubriendo fósiles de oso. El espeleotema ha proporcionado una datación de ca 70 ka.
LU-9. Similar a LU-7, pero sin macrofósiles, originado por caída de sedimentos desde las chimeneas. Espesor muy irregular.
Unidad aloestratigráfica AU-4.
LU-8. Colada de 2 cm de espesor.
LU-7. (Café Con Leche Clay o CCLC, debido a su color marrón claro). Espesor variable, alcanzando 150 cm. Limo arenoso, compuesto esencialmente por cuarzo, calcita, filosilicatos, principalmente illita, y una baja proporción de feldespato (plagioclasa y feldespato potásico). Las partículas de grano grueso incluyen bloques de piedra caliza desprendidos de las paredes y el techo, guijarros muy redondeados de piedra caliza y dolomita, fragmentos de guijarro cimentado, fragmentos de espeleotemas, margas centimétricas y clastos de arcilla roja, una gran acumulación de fósiles de carnívoros, principalmente U. deningeri y unos pocos fósiles de Hominini de pequeño tamaño. Los huesos y clastos están flotando, ya sea en la matriz o en contacto unos con otros y la distribución no es homogénea. Los guijarros de piedra caliza y dolomita y los fragmentos de conglomerado son alóctonos. Los sedimentos alóctonos se han datado por pIR-IR y TT-OSL en 428 ± 13 ka y 416 ± 17 ka, respectivamente (Arnold et al, 2014).
ES-3. Superficie erosiva muy irregular acompañada por un hundimiento en la zona central, limitado por pequeñas fallas normales en el depósito de arena. Presencia de grandes intraclastos incorporados de las unidades inferiores.
Unidad aloestratigráfica AU-3. Se forma a partir de cuatro procesos alternos: el goteo de agua con decantación de arcilla en todo el conducto, flujos de lodo procedentes de la rampa y flujos de agua que erosionan las unidades subyacentes, formación de cristales en balsas de agua no perturbadas y caídas del lecho rocoso y la cornisa.
LU-6 (Upper Red Clay o URC). Arcillas rojas con más calcita que en LU-5. Adoquines de diferentes litologías, derivados de estratos más antiguos. Alta densidad de fósiles de carnívoros y algunos fósiles de Hominini en contacto. Espesor muy irregular entre 0-50 cm. Polaridad normal. Espeleotemas muy frágiles, en posición original, formados a partir de agua estancada: escamas muy finas de carbonato y cristales gruesos intersticiales. Es producto de la infiltración del suelo a través de fisuras en el sistema de cuevas y goteo de agua. La datación de uno de estos cristales por T-series arrojó una edad media de 434 + 36 / -24 ka.
LU-6 contenía unos 7.000 fósiles humanos, la mayor acumulación de fósiles de Hominini jamás encontrada en un sitio del Pleistoceno Medio pertenecientes a unos 28 individuos de ambos sexos y distintas edades de muerte.
Los restos fueron clasificados como Homo heidelbergensis. Un estudio reciente (Arsuaga et al, 2014) confirma que el paleodemo de la Sima de los Huesos pertenece al clado neandertal, aunque más primitivo. Un genoma mitocondrial casi completo recuperado de un fémur no está relacionado con el de los neandertales, sino con el de los denisovanos (Meyer et al, 2014). La distribución era caótica; los elementos más completos por lo general adyacentes a la pared norte; más de 100 fósiles de pequeño tamaño se recuperaron de los sedimentos de la rampa.
También aparecieron muchos fósiles de Ursus deningeri y un pequeño número de otros taxones de carnívoros así como restos de microfauna.
Se halló una única herramienta de piedra: un bifaz.
Entre LU-6 y LU-5 no hay superficie erosiva significativa.
LU-5 (Lower Red Clay o LRC). Arcillas rojas con más cuarzo que en LU-6. Fósiles de Hominini y oso inconsecuentes con la unidad litoestratigráfica. Espesor máximo 50 cm. Huellas de polarización invertida en algunas muestras
ES-2. Erosión por corriente de agua, sin deposición asociada.
Unidad aloestratigráfica AU-2.
LU-4. Espeleotema. 3-6 cm de espesor.
LU-3. Sedimento detrítico. Depósito de arena fluvio-cárstico alogénico con un máximo espesor de 1,2 m. Estéril. Polaridad invertida.
ES-1. Superficie de erosión.
Unidad aloestratigráfica AU-1.
LU-2, cristalino grueso de 10 cm de espesor. Espeleotema datado en 856 + 110-65 ka (Arsuaga et al, 2014). Resedimentación a partir de la sala de los Cíclopes de origen fluvial y polaridad inversa.
LU-1, la unidad más baja, comprende margas blancas con ostrácodos del Mioceno Medio.
Secuencia estratigráfica muy bien conservada de más de 9 m, con evidencias de la prehistoria reciente (10 ka).
Galería de las Estatuas.
Se ha excavado en dos sectores distintos que corresponden al Pleistoceno Superior, en un momento en el que la sierra de Atapuerca estaba habitada por Neandertales. De los restos de macrofauna, los más abundantes son los de équido (familia de los caballos), los de ciervo y de bisonte. Algunos de estos restos han sido fracturados en fresco para la extracción de médula y también muestran marcas de corte. La cueva fue también usada por carnívoros de manera esporádica y se recuperaron restos de hiena y de zorro, así como algunos restos de ungulados con marcas de mordisco de estos carnívoros, que carroñeaban los restos dejados atrás por los neandertales.
Niveles de hasta 50 ka.
En 2017, entre los nivles 3 y 4 de GE-I (112-107 ka) se recuperó GE-1573, una falange del pie neandertal, junto con herramientas fabricadas en un amplio espectro de materiales, desde sílex a cuarcitas de todo tipo, en el que están representados lascas en bruto, raederas, denticulados y alguna punta, típicas de los neandertales.
En el extremo meridional de la Sierra de Atapuerca, a 1.033 m sobre el nivel del mar, se abre la Cueva del Mirador. Contiene 14 m de depósitos del último Pleistoceno y 6 m del Holoceno.
Una prospección arqueológica entre 1999-2003 descubrió 180 asentamientos al aire libre, 30 de ellos datados en MIS 4-3, la cronología correspondiente a los neandertales.
Sección de la Sima de los Huesos y esquemas de la formación de las unidades aloestratigráficas 3 (A) y 4 (B). Aranburu, Arsuaga y Sala, 2015.
Excavaciones y Estudios
1964. F. Jordá inicia la primera campaña de excavación en la Sierra.
1965. El Grupo Espeleológico Edelweiss (GEE) inicia una nueva topografía de las cavidades y enlaza la Cueva del Silo y la Cueva Mayor.
1966. Campaña de excavaciones dirigidas por Jordá, con el apoyo del GEE.
1972. El GEE descubre la Galería del Sílex.
1973. J. M. Apellániz excava el yacimiento del Portalón de Cueva Mayor, en la primera de las once campañas que realizaría (hasta 1983). Fruto de estos trabajos, con la colaboración del GEE, son dos monografías sobre la Galería del Sílex (1976 y 1987).
1976. T. Torres comienza a excavar en la Trinchera y en la Sima de los Huesos. Este último yacimiento proporciona el primer resto humano descubierto en Atapuerca, la mandíbula AT-1.
1977-1980. Replanteamiento topográfico de Cueva Mayor-Cueva del Silo.
1978. Unas voladuras de maniobras militares permiten el descubrimiento de la Cueva de los Zarpazos.
1979. Nueva topografía de la Galería del Sílex.
1980. Se inician las excavaciones en Galería. Las primeras intervenciones habían sido efectuadas en 1976.
1981. Se publica un estudio de todas las cavidades de la Sierra de Atapuerca (Martín et al).
1981-1989. Primeras intervenciones arqueológicas sistemáticas en Gran Dolina, afectando a una superficie de excavación de 30 m2 de TD10. A finales de los ochenta, en una prueba militar de explosivos, se perdieron parte de los sedimentos de los niveles TD4, TD5 y TD6.
1982-1995. Excavaciones sistemáticas en el yacimiento Galería.
1983. Emiliano Aguirre plantea las bases para el futuro de Atapuerca en un artículo.
1983. Estudio geomorfológico de la Sierra de Atapuerca (Zazo et al.).
1984. Se inician las excavaciones sistemáticas en la Sima de los Huesos.
1986. Primeras Intervenciones arqueopaleontológicas en la Sima del Elefante. Muestreo. Emiliano Aguirre.
1987. Primera monografía sobre Atapuerca (Aguirre et al.).
1990. El profesor y propulsor de las investigaciones y excavaciones de los yacimientos de la Sierra de Atapuerca, Emiliano Aguirre, se jubila y es sustituido por J.L. Arsuaga, J.M. Bermúdez de Castro y E. Carbonell, directores de las excavaciones hasta la actualidad.
1990-1991 Se excava el nivel TD4-3, el más antiguo de Gran Dolina con actividad antrópica. Se recogieron restos de caballos, ciervos, osos y rinocerontes, así como cuatro herramientas de cuarcita muy toscas.
1991. Aparecen instrumentos líticos Modo 1, junto a gran cantidad de fósiles de animales, en el nivel TD4 en la base de la Gran Dolina, datados en unos 900 ka.
1992. Se recuperan tres cráneos de la Sima de los Huesos. En Gran Dolina, se levanta el techo de la cueva y quedó expuesto a la intemperie TD11. Se colocó por ello una enorme cubierta.
1993. Primera publicación sobre Atapuerca en revistas internacionales de primer nivel (Arsuaga et al.).
1993. Sondeo de 9 m2 en Gran Dolina confirmando la Presencia de Materiales paleoarqueológicos en Todos Los Niveles Salvo TD2-1.
1994. Comienzo de la Excavación Sistemática de la Sima del Elefante.
1994-1995. En Gran Dolina aparecen fósiles humanos junto con abundantes herramientas líticas Modo 1 en el nivel TD6, datados en unos 800 ka.
1996. Se inician las excavaciones en la Sima del Elefante.
1996. Se inicia la excavación en extensión de Gran Dolina afectando una una superficie de más de 95 m2.
1996. Simposio Atapuerca y la Evolución Humana en la Fundación Ramón Areces.
1997. El Equipo Investigador de Atapuerca (EIA) define la especie Homo antecessor (Science; Bermúdez de Castro et al.). Monografía sobre la Sima de los Huesos en Journal of Human Evolution (Arsuaga et al.).
1998. Se encuentra un bifaz en la Sima de los Huesos.
1998. Primera monografía divulgativa sobre Atapuerca (Cervera et al.).
1999. Se inicia la excavación de la Cueva del Mirador.
1999. Monografía en Journal of Human Evolution sobre TD6 de Gran Dolina (Bermúdez de Castro et al.) y Trinchera-Galería (Carbonell et al.).
1999. Exposición Atapuerca, nuestros antecesores en el Museo Nacional de Ciencias Naturales de Madrid.
2000. Se recupera una herramienta lítica de los niveles inferiores de la Sima del Elefante.
2000. Empiezan las excavaciones en el yacimiento del Portalón de Cueva Mayor.
2001. Se define la especie Ursus dolinensis.
2001. Monografía en L'Antropologíe (Carbonell et al.).
2002-2010. Excavaciones en el yacimiento Galería.
2003. Se encuentran nuevos fósiles de Homo antecessor en TD6.
2003. Exposición en el Museo Americano de Historia Natural de Nueva York.
2004. Tesis doctoral de Alfonso Benito sobre la geomorfología y los paisajes neógenos y cuaternarios de la Sierra de Atapuerca y Valle Medio del río Arlanzón.
2006. Publicación divulgativa sobre Atapuerca (Arsuaga et al.).
2007. Se descubren restos de Homo antecessor datados en más de 1,2 Ma. Portada en Nature (Carbonell et al., 2008).
2008. Publicaciones de Ortega sobre el Portalón de Cueva Mayor.
2009. Tesis doctoral de Ana Isabel Ortega sobre la evolución geomorfológica del karst de Atapuerca.
2010
Sima del Elefante. Excavación en la zona central y norte. Fauna correspondiente a un paisaje de zonas arbóreas con otras algo más abiertas. Hallazgo de dos piezas líticas y huesos con marcas de corte en sedimentos por debajo
de 1,2 Ma.
Gran Dolina. Comienza la excavación del nivel TD6-3 y continúa la del nivel TD10-2. En este último se recuperan miles de restos faunísticos y de industria lítica, que ponen en evidencia una especialización en la explotación del bisonte con herramientas de sílex.
Galería de las Estatuas. Se amplían las áreas de excavación y se recuperan restos de fauna e industria lítica musteriense entre 50-45 ka.
Sima de los Huesos. Se termina de recuperar un cráneo cuyos primeros fragmentos se hallaron en 2009.
Hotel California. Se excavan cinco niveles arqueológicos del Paleolítico Medio y se da por clausurada la excavación comenzada en 2006 en la que se recuperaron cerca de un millar de
herramientas de piedra, realizadas por neandertales.
2011
Sima del Elefante. Se llega a la base del nivel TE9c.
Galería. Se reanudan los trabajos tras quince años de inactividad. Se interviene en la unidad GIV de la que se recuperan restos faunísticos y herramientas de sílex.
Gran Dolina. Finaliza la excavación de TD6 y comienza la excavación de TD5, en el que se recuperan restos de microfauna que sugieren un ambiente fresco y húmedo en el exterior y huesos de osos. Continúa la excavación de TD10-2, en el que se recuperan de nuevo miles de restos de fauna y líticos que confirman la especialización humana en el bisonte.
Galería de las Estatuas. Se recupera fauna e industria lítica musteriense.
Sima de los Huesos. Se halla un gran número de fósiles de oso y fragmentos humanos, entre los que destaca un fragmento de fémur.
2012
Sima del Elefante. Continúa la excavación de TE9, con la recuperación de restos de fauna.
Gran Dolina. Se excava TD4, donde aparece un cráneo de oso y un núcleo de sílex. En TD10, un bifaz de sílex y otro de cuarcita.
Galería de las Estatuas. Se recupera fauna e industria lítica musteriense.
Sima de los Huesos. Se recuperan varios restos humanos: un fragmento del hueso occipital del cráneo, gran parte de un húmero y una mandíbula (con muchos de sus dientes) y una tercera falange del dedo meñique del pie.
Fuente Mudarra. Se inician las excavaciones.
2013
Sima del Elefante. Se recupera una pieza de industria lítica en sílex cretácico del Modo 1 en el nivel TE8. Este nivel esta cerca del límite aceptado para la primera ocupación de Europa Occidental que se sitúa en 1,5 Ma.
Gran Dolina. En TD6, se termina de liberar un escápula infantil hallada en 2005. Se termina la excavación de TD10.2. En TD4 se recupera industria lítia que confirma la ocupación humana hace 1 Ma.
Galería. Se excava el subnivel GSU-3, datado entre 0,24-0,22 Ma, recuperándose restos faunísticos y algunas herramientas en sílex y arenisca correspondientes a las últimas ocupaciones de Galería.
En Sima de los Huesos y Galería de las Estatuas se realizan estudios geológicos y topográficos.
Fuente Mudarra. Se excava en un nivel de hace 50 ka, del que se recuperan herramientas líticas.
2014
Sima del Elefante. Se excavan TE9c y TE9d, recuperándose restos de fauna con señales de intervención antrópica. En TE9d (más antiguo que 1,2 Ma) aparece una lasca de industria lítica en sílex neógeno.
Gran Dolina. Se excava el techo del nivel TD10.3, de unos 420.000 años de antigüedad, en el que aparecen unos pocos instrumentos líticos y fósiles de caballo. En TD4 se descubren algunas herramientas de sílex que seguramente sirvieron para procesar las carcasas de los animales atrapados en el foso. Un análisis paleomagnético indica que desde el nivel TD7 hacia abajo, todos los sedimentos son más antiguos de 780 ka.
Galería. La unidad GIIlb, datada en 260 ka, ha proporcionado más de 500 restos de fauna y más de 40 restos líticos: lascas de sílex neógeno, utilizadas para descarnar a los animales, y bases naturales de cuarcita, usadas como percutores para romper huesos y/o retocar las herramientas. Los hallazgos sugieren que los Homo entraron en Galería de manera esporádica para aprovechar a los animales que caían a través a de la trampa natural situada al Este de la cavidad. La actividad de carnívoros también ha sido identificada a través de mordeduras en los restos abandonados por los humanos.
Sima de los Huesos. Se hallaron nuevos fósiles humanos.
Galería de las Estatuas. Se recuperaron restos de fauna y piezas de industria musteriense. La cronología de los niveles superiores se sitúa en torno a los 50-45 ka.
Fuente Mudarra. En el nivel, 4b se recuperan cientos de restos que indican que en este lugar se realizaban labores de manufactura de instrumentos líticos de sílex y cuarcita hace 50 ka.
2015
Sima del Elefante. Se excavó la base del nivel TE9c y se destapó TE9d en prácticamente toda la superficie de excavación. Este nivel contenía restos arqueopaloentológicos de gamos o ciervo gigante, que en algunos casos presentaban marcas de corte producidas por los Homo. Se efectuó un sondeo bioestratigráfico (de unos dos metros cuadrados) en el nivel TE7 con el fin de conocer la potencia del paquete sedimentario, comprobando su riqueza paleontológica, como se aprecia con los restos de ciervo gigante e hipopótamo, así como con los abundantes restos de micromamíferos.
Gran Dolina. Se trabajó en el tramo medio del nivel TD10.3, de unos 0,45 Ma de antigüedad, documentando una menor concentración de restos con evidencias de actividad humana que en las capas más altas. Han aparecido pocos instrumentos líticos, y los huesos con marcas de procesado por parte de Homo son escasos. Destaca la alta representación de grandes carnívoros. En TD4 se excavaron 15 m2 y se recuperaron algunas herramientas líticas junto con restos de fauna.
Galería. Se excavó Unidad GIIIa, datada en 0,27 Ma, en toda la superficie intervenida, proporcionando abundantes restos de fauna, principalmente de caballos y ciervos pero también de carnívoros, y algunas herramientas líticas.
Sima de los Huesos. Finalizó la excavación del área AU-3, recuperando nuevos e importantes fragmentos craneales, corroborando la cronología avanzando en la localización de la antigua entrada a la cavidad.
Galería de las Estatuas. Se han recuperado restos líticos y de fauna cazada, principalmente équidos y ciervo, en muchos casos con marcas de corte. Hay evidencias de que los carnívoros, como la hiena manchada, también utilizaron la cavidad de manera esporádica.
Fuente Mudarra. Se trabajó en el Nivel 4B, datado en hace 56 ka, recuperando herramientas de sílex y cuarcita, sobre todo restos de la fabricación de cuchillos, raederas y denticulados. Las huellas de uso estudiadas en los filos de estos útiles nos demuestran que los utilizaban con piel, carne y hueso.
Se iniciaron prospecciones mediante georradar (GR) y tomografía de resistividad eléctrica (TRE) en zonas próximas a la Trinchera del Ferrocarril para detectar posibles yacimientos. En la Cueva del Fantasma se ha procedido a la detección de posibles rellenos fosilíferos. Se realizó un sondeo mecánico en el lugar señalado por los análisis de GR y TRE para la posición de la cavidad que conecta las cuevas de la Gran Dolina y Covacha de los Zarpazos.
2016
Sima del Elefante. Finalizó la excavación del nivel TE9d, con una cronología de ca 1,3 Ma, recuperando restos de aves de diferentes tamaños, oso, bóvido y ciervo. Algunos restos de herbívoro presentan marcas de corte y fracturación de los huesos largos. También se ha recuperado un fragmento de lasca en sílex cretácico. En la excavación de TE9d, se ha alcanzado el nivel TE7, que es el más antiguo de toda la Trinchera del Ferrocarril y del que solo se tenía evidencia en la zona sur del yacimiento.
Gran Dolina. Se ha trabajado en el tramo inferior del nivel TD10.3, de unos 450 ka de antigüedad. Apareció una menor concentración de restos que en las capas superiores, con representación de diversas especies de herbívoros y carnívoros. La actividad antrópica sobre estos restos es escasa. Está bien representada la cultura Achelense, con sus característicos bifaces y otros grandes instrumentos, aunque, en consonancia con la escasa actividad antrópica sobre los restos de fauna, la densidad de instrumentos es baja. Las excavaciones en TD4 permiten reafirmar la presencia humana. Entre los restos más destacables, aparte de la industria lítica, destacan multitud de astas de desmogue de diversos cérvidos y dos mandíbulas completas de jaguar europeo.
Galería. Se excavaron los niveles inferiores de la Unidad GIIIa, datada entre 280-220 ka, que ha proporcionado abundantes restos de fauna e industria lítica. Las herramientas son características del Modo 2 (Achelense). Se han recuperado lascas, bifaces y hendedores elaborados en distinta materia prima como es el sílex, la arenisca y la cuarcita. Los restos faunísticos recuperados (costillas, vértebras y elementos craneales) pertenecen principalmente a caballos, habiendo recuperado también elementos esqueléticos de otros herbívoros y carnívoros. Todo apunta a un aprovechamiento antrópico de los animales que caían por la trampa natural situada al Este de la cueva. Los grupos humanos del Pleistoceno Medio de la sierra de Atapuerca accedían a los animales y transportaban los elementos esqueléticos más ricos en masa muscular (extremidades), abandonando en Galería los restos pertenecientes a la caja torácica. Los huesos abandonados eran, posteriormente, aprovechados por los carnívoros, como lo atestiguan las múltiples mordeduras halladas en los restos.
Sima de los Huesos. Se ha trabajado en la delimitación del contacto entre las unidades estratigráficas 6 y 7 en la zona central del yacimiento. La unidad 6 contiene fósiles humanos y de carnívoros (mayoritariamente osos) y la 7 solo de carnívoros. El volumen de sedimento excavado de la unidad 6 es todavía limitado. No obstante, se han recuperado algunos fósiles humanos, que incluyen tres falanges, un extremo de tibia, otro de peroné y un tercero de húmero, parte de una clavícula y algunos fragmentos de costillas y de cráneo.
Galería de las Estatuas. Se ha profundizado en las dos áreas de excavación y se han recuperado restos de fauna y piezas de industria musteriense. Los restos de fauna incluyen équidos y lepóridos consumidos por los neandertales. La cronología de los niveles superiores se sitúa en hace ca 52-48 ka, y los estudios del polen indican que se produjo un cambio de paisaje, pasando de ser un ambiente abierto de tipo estepa a bosque abierto.
Fuente Mudarra. Se ha continuado excavando el nivel 4, muy rico en herramientas de piedra fabricadas por los neandertales. Por debajo del Nivel 4 se ha encontrado otro nivel en el que se está trabajando y en donde ya se han recuperado cientos de piezas de sílex con para descuartizar y tratar las pieles de diferentes animales.
La campaña de prospecciones geofísicas se ha centrado en el sector de Torcas, con el fin de conocer la morfología de las cavidades de Trinchera del Ferrocarril, con especial detalle de los yacimientos Gran Dolina, Galería y Elefante. Las prospecciones mediante la técnica de tomografía de resistividad eléctrica (TRE) se han desarrollado en el término de Torcas, con el objetivo de investigar el desarrollo de las cavidades del yacimiento de la Sima del Elefante y los conductos de Cueva Mayor y Cueva Peluda. Estas prospecciones han permitido conocer la presencia de nuevas cavidades, así como determinar el desarrollo y la posición de las antiguas entradas. Un conjunto de sondeos mecánicos se ha realizado en los lugares señalados por los análisis de GPR y TRE de la campaña de 2015 en la ladera de Torcas, confirmando la presencia del conducto de Dolina y Galería, con una sección de unos 9m, o la presencia de una nueva cavidad en cotas del primer piso.
2017
Sima del Elefante. Se excavó TE7, sin indicios de ocupación humana.
Gran Dolina. La excavación de la parte inferior de TD10 proporcionó una gran acumulación de industria lítica, en la que destaca una proporción inusual de bifaces.
Galería. Se ha excavado la parte inferior de la Unidad GIIIa. En este tramo de la secuencia, datado en hace ca 270 ka, se han recuperado restos de fauna e industria lítica principalmente en sílex. También se han recuperado cantos de cuarcita que fueron utilizados como percutores o machacadores de huesos. Uno de los principales hallazgos corresponde a un núcleo de sílex neógeno de grandes dimensiones. Éste fue introducido intencionalmente en la cueva por los humanos como remanente de materia prima para producir lascas que les permitiesen procesar las carcasas de los animales caídos por la trampa natural en sucesivas visitas a la cueva. De este modo se reafirma el uso reiterado de la cavidad como lugar de obtención de comida.
Sima de los Huesos. Se recuperó la mitad de una mandíbula humana en una zona todavía no explorada.
Galería de las Estatuas. Se ha excavado en dos sectores distintos que corresponden al Pleistoceno Superior, momento en que la sierra de Atapuerca estaba habitada por Neandertales. En esta campaña se han recuperado una falange humana, restos de fauna e industria lítica. Se trata del primer resto neandertal localizado en Atapuerca, a la espera de confirmar si también lo es un parietal encontrado en 2016 en la Cueva Fantasma. De los restos de macrofauna, los más abundantes son los de équido y los de ciervo. Algunos de estos restos han sido fracturados en fresco para la extracción de médula y también muestran marcas de corte. En esta campaña también se ha recuperado un resto de lagomorfo con marcas de corte. La cueva fue también usada por carnívoros de forma esporádica. Este año se han recuperado restos de hiena, y por primera vez en este yacimiento, de lobo. Las herramientas de piedra recuperadas están hechas en dos tipos distintos de sílex, y destaca la presencia de raederas. También se ha recuperado un fragmento de diáfisis de un hueso largo de fauna que fue utilizado como retocador.
Fuente Mudarra. En el nivel 8, se ha recuperado un rico conjunto de sílex, cuarcita y cuarzo entre los que caben destacar percutores para golpear los nódulos de sílex, y las lascas y cuchillos resultantes, que se usaron unas veces allí mismo para descuartizar animales y trabajar madera, y otras se llevaron a otros lugares de la Sierra.
Cueva Fantasma. La retirada de unas 12.000 toneladas de roca del techo derrumbado de la cueva ha terminado por mostrarnos una superficie de unos 300 metros cuadrados. Los sondeos con testigo continuo y las prospecciones geofísicas llegan hasta los 15 metros de profundidad. En 2016, durante las labores de limpieza, apareció un parietal humano.
2018
Sima del Elefante: Se han
hallado diferentes restos de animales entre los que destacan el pigargo y el
castor, los cuales “apuntan a la existencia de un ambiente húmedo en la Sierra
hace 1,3 Ma”, junto con ciervo gigante y gamo. Aunque, hasta el momento, en el
nivel 7 no se han obtenido evidencias directas o indirectas de presencia humana,
el equipo “se encuentra en la fase inicial de excavación y es posible que en
futuras campañas aparezcan”. En TE9 han aparecido las mandíbulas de las musarañas gigantes muy bien conservadas que indican que en esos momentos las condiciones de humedad y temperatura favorecieron un paisaje rico y con agua abundante en los alrededores de la Cueva.
Gran Dolina: El nivel TD10.4, datado en algo más de 400 ka, algo más pobre que los superiores, ha aportado una significativa muestra de instrumentos de piedra, entre los que destacan elementos característicos del Achelense como bifaces y hendedores. En cuanto a la fauna, mal conservada debido a las condiciones ácidas del estrato sobre el que apoya, destaca la presencia tanto de ungulados como carnívoros y, como novedad, la de oso. También el molar de un gran erizo, que indica que en ese tiempo el clima era templado y húmedo, con vegetación y alimentación abundante. Al concluir esta campaña se ha puesto al descubierto el techo del nivel TD9 en toda la superficie. Los numerosos fósiles de oso que se han recuperado este año en el nivel TD4 de la Gran Dolina pertenecen a la especie Ursus dolinensis.
Galería: La campaña ha permitido alcanzar la base de la Unidad GII caracterizada por abundantes restos de fauna e industria lítica de una antigüedad de unos 300 ka. Principalmente, se han localizado restos de caballos y ciervos (huesos axiales y craneales) asociados a bifaces y a núcleos utilizados para extraer lascas con las que procesar a estos animales. Se han documentado además abundantes cantos de río utilizados como machacadores de huesos y percutores para la talla. También se han recuperado restos de carnívoros, entre los que destaca una hemimandíbula derecha de león en perfecto estado de conservación.
La Paredeja. Con el fin de evaluar el potencial arqueológico de este relleno de nueve metros de espesor, se ha procedido a limpiar el corte escalonando el terreno. Se han identificado diferentes unidades estratigráficas con varios niveles arqueológicos que contienen herramientas de piedra fabricadas con sílex y cuarcita, así como algún resto de fauna en los niveles inferiores. A la espera de dataciones y analizando las herramientas de piedra, todo indica que se trata de un rico asentamiento ocupado durante el paleolítico medio (150-30 ka).
Sima de los Huesos: Se ha extendido la excavación en la zona noreste del yacimiento sobre la que aún no se había actuado. Se recuperaron una veintena de fósiles humanos que incluyen el lado izquierdo que faltaba del cráneo número 16 (Sara).
Galería de las Estatuas: Se han recuperado fósiles de macromamíferos (ciervos, équidos, hienas) así como de aves y restos de industria lítica. Además también se ha avanzado en la toma de muestras para caracterizar mejor las cronologías de estas ocupaciones.
Cueva Fantasma: Después que durante los últimos dos años se procediera a la limpieza de la vieja cantera, este año se ha comenzado a excavar en este yacimiento. La primera fase de las labores se ha centrado en un sondeo del sector del conducto o cavidad ubicada al oeste de Cueva Fantasma, para continuar con la excavación en sí misma. Se ha alcanzado casi el metro y medio de profundidad donde se han encontrado numerosos restos fechables durante el Pleistoceno Superior, es decir, de una antigüedad de unos 50 ka. Entre ellos, restos de caballo, de otros bovinos, de mustélidos y algunos pequeños carnívoros como el Cuan (perro salvaje).
2019
Sima del Elefante: Desde 2012 se estaba excavando en dos áreas separadas, sin continuidad sedimentaria entre ellas. En la campaña de 2019 las dos áreas se han unido en la superficie del nivel 7, siendo este el único nivel que han excavado hasta el momento en toda la superficie del yacimiento, con una cronología alrededor de 1,4 Ma, con la recuperación de los restos de un individuo inmaduro y las extremidades superiores de un adulto en conexión anatómica, así como restos de fauna que confirman un ambiente templado y húmedo.
Gran Dolina: Se ha terminado la excavación del nivel TD10.4, datado en algo más de hace 400 ka. A continuación, se ha excavado el nivel TD9 donde las características del sedimento han impedido la conservación de restos de fauna y solo se han recuperado algunos instrumentos líticos. Se ha dejado al descubierto el techo de la unidad TD8, cubierto prácticamente en su totalidad por formaciones estalagmíticas, características de un ambiente de cueva cerrada. La excavación del nivel TD4, datado en cerca de un 1 Ma ha permitido la recuperación de nuevos restos de Ursus dolinensis así como de cérvidos de cinco especies.
Galería: La excavación ha alcanzado la base de la Unidad GIIIa, correspondiente al nivel TG10A, con alrededor de 300-280 ka de antigüedad. Los restos faunísticos pertenecen principalmente a costillas, vértebras y mandíbulas de caballos y ciervos, que cayeron por la trampa natural y fueron procesados por los humanos, como testimonian las marcas de corte y fracturas intencionales. La industria Achelense localizada en el yacimiento se caracteriza por la presencia de bifaces y pequeñas lascas retocadas (puntas y raederas) y están elaboradas mayoritariamente en cuarcita y sílex neógeno. También son abundantes los cantos de río aportados por los humanos como percutores o machacadores de huesos. Con todas estas herramientas los humanos preparaban a los esqueletos caídos para ser transportados a sus lugares referenciales.
Cueva Fantasma: Ha continuado el sondeo empezado en la campaña anterior (2018), localizado en el sector de la antigua entrada de esta cavidad. La recuperación en los niveles 28d y 27 de dos lascas de tecnología de Musteriense evidencia la presencia de actividad humana, en una fase de la secuencia estratigráfica posiblemente relacionada con MIS 3. También ha continuado la excavación en extensión de la zona interna, o Sala Fantasma, formada por un espacio de unos 300 m2. La intervención ha puesto al descubierto una gran cantidad de restos arqueo-paleontológicos, que cubren la mayor parte de la superficie de la sala, entre los que destaca el predominio de restos de équidos, con prácticamente representación de todo el esqueleto. Junto a los caballos también hay bóvidos, cérvidos y suidos. Entre los carnívoros se han recuperado restos de hienas, osos, cánidos (lobo y zorro), tejón, león y lince. Este registro faunístico posiblemente esté relacionado con cubiles de hienas y otros carnívoros, aún sin datar. La superficie excavada posiblemente se encontrase muy cerca del techo de la cavidad, y por lo tanto presentaba un tamaño reducido apto para la ocupación de estos animales. Sin embargo, durante la intervención en extensión en la Sala Fantasma también se ha registrado una pieza de industria lítica (un cuchillo de dorso de tipología musteriense), que evidencia el uso de este espacio por los neandertales.
Sima de los Huesos: Han proseguido las excavaciones en la zona norte y la parte central, con la recuperación de una decena de fósiles humanos entre los que destaca un fragmento de pelvis, que ayudará a enteder la dinámica del parto y la paleobiología de aquellos humanos. También cabe destacar un hueso temporal que probablemente corresponda a alguno de los cráneos descubiertos en campañas anteriores.
Galería de las Estatuas: Se ha excavado en dos sectores distintos que corresponden al Pleistoceno Superior, en un momento en el que la sierra de Atapuerca estaba habitada por Neandertales. En la presente campaña se han recuperado tanto restos de fauna como herramientas de piedra. De los restos de macrofauna, los más abundantes son los de équido (familia de los caballos), los de ciervo y de bisonte. Algunos de estos restos han sido fracturados en fresco para la extracción de médula y también muestran marcas de corte. Esta campaña se ha recuperado un resto de costilla que preserva una marca de corte con restos de la herramienta que la produjo incrustada. La cueva fue también usada por carnívoros de manera esporádica. Este año se han recuperado restos de hiena y de zorro, así como algunos restos de ungulados con marcas de mordisco de estos carnívoros, que carroñeaban los restos dejados atrás por los Neandertales. Por último, mencionar que durante esta campaña también se han recuperado varios restos de puercoespín, especie poco abundante durante el Pleistoceno Superior. Las herramientas de piedra recuperadas están hechas tanto de sílex como de cuarcita.
La Paredeja: En el nivel arqueológico superior, se ha recuperado un rico conjunto de herramientas de sílex y cuarcita realizadas por grupos de neandertales. Se ha iniciado un sondeo para conocer los posibles niveles arqueológicos que puede haber por debajo del nivel del suelo actual hasta llegar a la base de la cavidad.
Sima del Elefante: Se excavó parte del nivel TE7, eliminando grandes bloques caídos del techo. Se recuperaron restos de tortuga, oso y castor, lo que refuerza la hipótesis de que hace 1,4 Ma el clima era más templado y húmedo que en la actualidad.
Gran Dolina: En el remate de la excavación de TD9 aparecieron herramientas líticas. Comenzó la excavación de TD8 con abundantes restos de macromamíferos, entre los que destacan animales como el oso, la hiena o el jaguar, además de herbívoros como el rinoceronte, el caballo o los cérvidos. La gran novedad de esta campaña ha sido la aparición, junto a esta fauna, de dos utensilios de cuarcita: una lasca y un canto tallado. Por lo tanto, se documenta por primera vez la presencia humana en la Sierra en una franja temporal, entre los 600-500 ka, de la que no había evidencia en ninguna de las cavidades de la sierra de Atapuerca.
Cueva Fantasma: La excavación de 2020 ha continuado con la excavación del nivel 27c del Sector Entrada, sin apenas restos arqueológicos, alcanzando el nivel 26 en toda la superficie, descubriendo cierta concentración de elementos arqueo-paleontológicos de équidos, restos de zorros y de conejo. Este nivel 26 destaca por la presencia de industria, recuperando un conjunto de 6 piezas de tecnología de musteriense (entre las que destaca un cuchillo de dorso), que evidencian presencia de actividad humana y uso de este espacio por parte de los neandertales, en una fase de la secuencia estratigráfica posiblemente relacionada con MIS 3. En el perfilado del frente oriental de la de Sala Fantasma (PSF) se ha documentado la predominancia de restos de équidos, con prácticamente representación de todos los elementos del esqueleto, seguidos de restos de bóvidos, cérvidos y suidos. Entre los carnívoros se han recuperado restos de hienas, osos, cánidos (lobo y zorro), tejón y lince. Esta unidad ha proporcionado en el sector próximo a Cueva Fantasma varias lascas de sílex, que evidencia el uso de este espacio por parte de los Neandertales, siguiendo la pauta descrita en el sector de entrada de Cueva Fantasma. En la parte inferior del perfilado, destaca la unidad 21 por la abundancia de restos fósiles de hiena. Además se han recuperado un gran número de coprolitos de hiena, que indican el posible uso de este espacio como letrina. En este nivel además se han recuperados restos de équidos, bóvidos, cérvidos y rinocerontes. También se ha podido recuperar restos de otros grandes carnívoros como el león, pantera y oso. Por su parte la unidad 20, se caracteriza por un nivel cementado de conglomerados y bandas de arcillas que contienen restos fósiles de herbívoros (équidos, bóvidos, rinocerontes y especialmente ciervos) y carnívoros (pantera, hiena, osos).
Sima de los Huesos: Fragmento de hueso occipital.
Galería de las Estatuas: La gran novedad de este año es la excavación de la boca de entrada de la Galería de las Estatuas (yacimiento de Estatuas Exterior), que se cerró hace más de 50.000 años. En esta boca de cueva habitaron los neandertales. La industria lítica recuperada en el yacimiento de Estatuas Exterior está realizada en materias primas del entorno de la sierra de Atapuerca como el sílex, la cuarcita y la arenisca, presentando además una clara afinidad con el tecnocomplejo musteriense, que es el de los neandertales. Tanto las materias primas como la forma de tallar dichas herramientas muestran grandes similitudes con el conjunto lítico hallado en el yacimiento de Galería de las Estatuas, que es su continuación en el interior de la Cueva Mayor. En la Galería de las Estatuas Interior se ha excavado el nivel 5 de la cata Estatuas I, que es el más antiguo de la secuencia, con una edad superior a los 110 ka. Aunque este nivel 5 apenas contiene fósiles, se ha recuperado una mandíbula de puercoespín (Hystrix) y un coprolito de carnívoro. El puercoespín es un animal relativamente escaso en el registro del Pleistoceno final, habiendo sido localizado en menos de 10 yacimientos en toda la península ibérica. En el caso de Estatuas II, se ha continuado trabajando en el nivel 3, que ahora está expuesto en toda la cata y que corresponde al final del último periodo interglacial. Se han recuperado algunas herramientas retocadas de sílex y varios restos fósiles.