 |
| Izquierda Sangiran 17. Derecha Broken Hill Homo rhodesiensis |
Dubois descubrió cerca de Trinil una calota en 1891 y un fémur en 1892, a partir de los cuales definió el Pithecanthropus erectus.
 |
| Niño de Mojokerto |
La presencia de estos Homo en Java plantea el problema de cómo cruzaron las aguas:
- Hipótesis de la desecación de ciertos mares en esa época. Java
está unida al continente por una plataforma continental poco profunda.
- Se han encontrado elementos líticos y al Homo floresiensis en la isla de Flores que se encuentra aislada
por aguas muy profundas. Si el Homo
erectus fue capaz de construir balsas hay que concederle una capacidad
cognitiva muy elevada. Los restos de Flores se han datado en 75 ka.
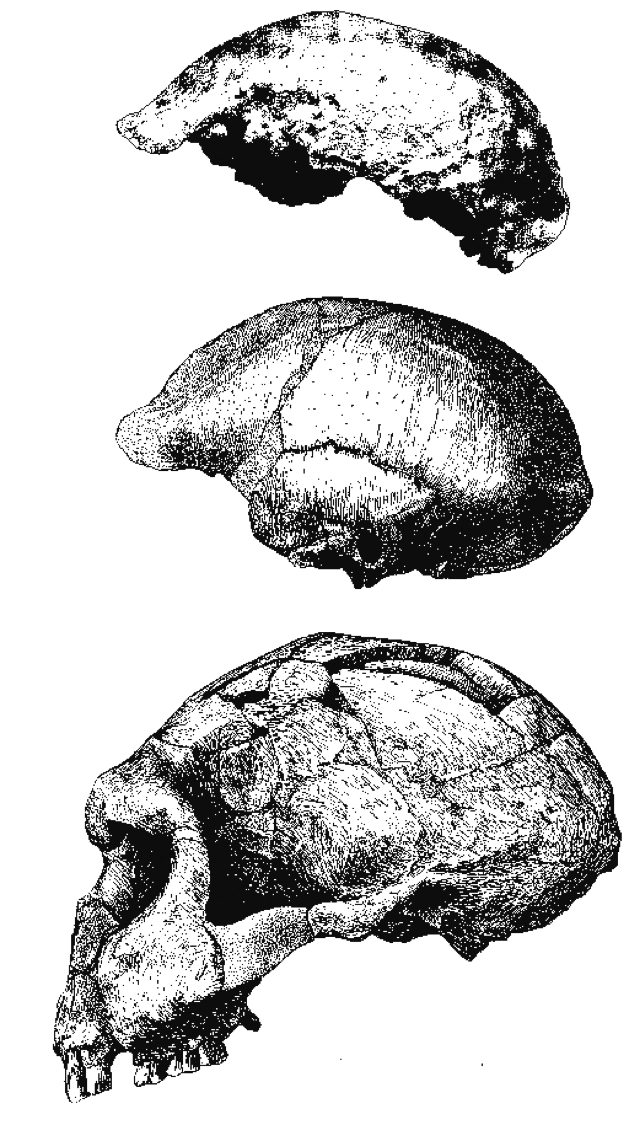 |
| De arriba a abajo: Trinil, Sangiran 2, Sangiran 17 |
Hallazgos:
Trinil- Dubois descubrió cerca de Trinil fósiles humanos en 1891-1892, a partir de los cuales definió el Pithecanthropus erectus.
- Java Man, Pithecanthropus I, (Pithecanthropus erectus). Descubierto por Eugene Dubois en 1891 cerca de Trinil (Java). Holotipo de Homo erecuts. Su edad es de menos de 1 Ma. Calota gruesa con un perfil muy bajo, frontal muy huidizo y torus supraorbitario relativamente poco marcado; cresta sagital; ancho máximo en la base del cráneo; pómulos planos, con marcada tuberosidad cigomaxilar; marcado borde inferior orbital; falta de demarcación entre la región nasal y la región inferior de la cara; toro occipital muy marcado en forma de barra continua. Capacidad craneana 940 cc. Atribuido al sexo femenino.
- El siguiente año se encontraron Trinil Fémur I, 12 metros abajo, de morfología similar a la de los humanos actuales, que pudo pertenecer a un individuo de 1,70 m y dos muelas.
- Trinil 3 (Fémur I). Fémur izquierdo completo pero patológico hallado en agosto de 1892. Para Ruff et al (2013, 2015) no corresponde con erectus y es más joven que el resto de la colección. Los niveles en el hueso de Ca y P indican que fue extraído de un diferente nivel estratigráfico (Bartsiokas y Day, 1993).
- Posteriormente se recuperaron otros cuatro fémures (Trinil Fémures II-V).
- Trinil 6 (Fémur II)
- Trinil 7 (Fémur III)
- Trinil 8 (Fémur IV)
- Trinil 9 (Fémur V)
- Josephine C. A. Joordens et al (2014) han informado de la existencia de un grabado geométrico (DUB1006-FL) en una concha de almeja de agua dulce de la colección Dubois (museo Naturalis, Leiden, Países Bajos) que reúne los hallazgos de Trinil. En esta colección se halló también evidencia del consumo de almejas de agua dulce (Pseudodon) utilizando dientes de tiburón para perforar la concha y cortar el músculo que las mantiene cerradas, y una herramienta de concha: una valva con signos de modificación por retoque (DUB5234-dL). Las ranuras en la concha DUB1006-FL, cada una de las cuales es de aproximadamente 1 cm de largo, muestran signos de envejecimiento significativo, y no hay espacios entre las vueltas, lo que indica que el grabador hubo de prestar atención a los detalles. El grabado probablemente se realizó en una cáscara fresca, con la mano diestra, utilizando un diente de tiburón, en una sola sesión, por una sola persona manteniendo firmemente la concha y la herramienta para cambiar de dirección. En aquel momento las líneas serían blancas sobre un fondo oscuro. En conjunto, los datos indican que el grabado fue realizado por el Homo erectus. Tanto el grabado como la forma de abrir las almejas, que requiere un alto nivel de destreza y conocimiento de la anatomía del músculo, sugieren un alto nivel de cognición y control neuromotor.
 |
| Cronología de fosiles asiáticos de Homo arcaico. Kaifu, 2017. |
Ngandong. Solo: investigadores holandeses descubrieron más fósiles en la década de 1930 cerca de la aldea de Ngandong, en el río Solo. Hasta hace poco, se pensaba que representaban una población muy reciente, pero el trabajo de campo y los análisis más recientes sugieren que aquellos Homo vivieron, en algún momento entre 143-546 ka. 11 calotas y 2 tibias. Los cráneos tienen las superestructuras muy marcadas y sus bóvedas son ligeramente más altas. Capacidades en torno a 1.150 cc. La morfología parece corresponder a un estadio evolutivo final del erectus. Datados en 0,05 Ma.
- Ngandong-6. 1.251 cc.
- Ngandong-7. 1.013 cc.
- Ngandong-10. 1.135 cc.
- Ngandong-11. 1.231 cc.
- Ngandong-12. 1.090 cc.
- En Ngandong y en la localidad próxima de Matar se hallaron lascas y algunos núcleos rudimentarios (Fauzi et al, 2016).
- El niño de Mojokerto es un cráneo infantil descubierto en Perning (Mojokerto) en 1936, de 630 cc. Torus supraorbitario incipiente. Las teorías sobre la edad a la muerte van desde 1 hasta 8 años.
 |
| Estratigrafía de Sangiran y lugar del hallazgo de Bpg 2001.04. Zaim et al, 2011. |
Algunas mándíbulas tiene un aspecto masivo, con dientes de aspecto primitivo y de gran tamaño (Kaifu et al, 2005, Kaifu, 2006). Estos fósiles han originado polémicas muy interesantes sobre la posibilidad de que los australopitecos abandonaran África hace más de dos millones de años y recalaran en el sudeste asiático antes de que lo hiciera el género Homo. No obstante, se ha llegado al consenso de incluir a todos ellos en la especie Homo erectus. Sangiran 2 fue el primer ejemplar recuperado (1937). Datados en 1,51 Ma. (Larick et al, 2001) - 900 ka. (Hyodo et al, 1993).
Los ejemplares más importantes son:
- Sangiran 2, Pithecanthropus II. Descubierto por G.H.R. von Koenigswald en 1937 en Sangiran. La bóveda craneal es muy similar a la del primer cráneo del Hombre de Java, pero menos completa y con una capacidad craneal inferior: sólo 813 cc. 1,5 Ma.
- Sangiran 4. 908 cc. 1,6 Ma.
- Sangiran 10. 855 cc. 1,2 Ma.
- Sangiran 12. 1.059 cc. 1,1 Ma.
- Sangiran 17, Pithecanthropus VIII. Descubierto por Sastrohamidjojo Sartono en 1969 en Sangiran. Se trata de un cráneo casi completo, con una capacidad craneal de 1.004 cc. Es el fósil de erectus más completo de Java. Este cráneo es muy robusto. Tienen la cara prominente, muy ancha y bastante plana y los pómulos pronunciados. Sin asociación a instrumentos líticos. 1,3 Ma.
- Sangiran 31. Descubierto en 1979. Parte posterior de un cráneo. Paredes muy gruesas y torus occipital muy marcado.
- En 2001, aparecó Bpg 2001.04, un fragmento maxilar (preservando P3-M2), en la Formación Bapang, entre gravas cementadas en la zona Grenzbank. Dos metros por encima del punto del hallazgo, una datación 40 Ar / 39 Ar ha producido una edad de 1,51 ± 0.08 Ma. Los patrones dentales sugieren una demo diferente a la representada por los anteriores hallazgos (Zaim et al, 2011).
- En 2016, se recuperó un cráneo adicional, de unos 800 cc.
- Diente humano junto con una industria lítica muy rica de Modo 1 y numerosos restos de fauna.
- La presencia lítica es una particularidad de este yacimiento ya que en Java los artefactos son raros.
- Sm-1 (1973). Calota asociada a dos restos líticos. 1.200 cc-
- Sm-2 (1977). Fragmento distal de diáfisis de tibia con morfología moderna.
- Sm-3 (1997). Cráneo. 917 cc.
- Sm-4 (2003). Cráneo. 1.007 cc.
Ngawi.
- Calota de individuo adulto.
Punung
- PU-198, tercer premolar maxilar, de tamaño relativamente pequeño, pero dentro del rango de variación de erectus y sapiens. Datado en hace 143-115 ka. Es preciso tener en cuenta la tendencia hacia la disminución del tamaño de los premolares entre los erectus de Java tempranos y tardíos (Polanski, Marsh y Maddux, 2015).
 |
| Ngandong 11 |
Interpretaciones
Los erectus asiáticos han sido considerados por algunos autores. los únicos erectus en sentido estricto. El niño de Mojokerto fue datado por Swisher en más de 1,8 Ma y eso significaría que la salida de África es anterior al achelense. Para él, el erectus evolucionó fuera de África (hipótesis de la salida temprana). Esta hipótesis ha sido puesta en duda por la dificultad de datación.
Para Emiliano Aguirre (2008) estos fósiles pertenecen a un grupo descendiente de unos seres que salieron de África en el momento de la cladogénesis del ergaster.
Unos ejemplares dentales muestran un carácter robusto y otros más grácil por lo que se ha especulado en un pronunciado dimorfismo sexual. Otra interpretación (Zanolli et al, 2019) justifica la denominación de Meganthtropus palaeojavanicus a los ejemplares más robustos, en concreto:
Para Emiliano Aguirre (2008) estos fósiles pertenecen a un grupo descendiente de unos seres que salieron de África en el momento de la cladogénesis del ergaster.
- Trinil 11620
- Trinil 11621
- Sangiran 5
- Sangiran 6a
- Arjuna 9
- FS-77
- SMF-8864
Las dataciones de estos ejemplares no son muy diferentes a las de Daminisi, pero sí lo es su morfología (François Sémah, 2014). La mayor parte de los hallazgos son similares al holotipo hallado por Dubois y se clasifican en la denominación Grupo Sangiran-Trinil (Grimaud-Hervé et al, 2012). Fueron recuperados de una capa de sedimentos volcánicos datada en 0,9-0,7 Ma (Kabuh layers).
Swisher et al (1994) dataron los restos de Mojokerto en 1,81 Ma y los de Sangiran en 1,6-1,1 Ma. En 2011, Russell L. Ciochon, Yahdi Zaim y E. Arthur Bettis dataron en 1,5 Ma unos restos de mandíbula hallados en Sangiran. Josephine C. A. Joordens et al (2014) dataron sedimentos contenidos conchas de mejillones de agua dulce de Trinil mediante 40 Ar / 39 Ar y luminiscencia, obteniendo una edad máxima de 0,54 ± 0,10 Ma y una edad mínima de 0,43 ± 0,05 Ma, lo que supone una fecha mucho más joven que la considerada hasta ahora.
- Los fósiles de finales del Pleistoceno Medio y Pleistoceno Tardío son más derivados y están representados por Solo, Ngandong, Sambungmacan y Ngawi (Homo soloensis) y en ellos destaca su encefalización (Grimaud-Hervé, 1997). A menudo son datados entre hace 70-30 ka (Antón 2003; Yokoyama et al 2008). Sin embargo, esta es una suposición injustificada, principalmente debido a la asociación estratigráfica incierta entre los materiales fechados y los restos humanos debido a que la acumulación de fósiles de grandes mamíferos en las terrazas fluviales fue producida por un ambiente deposicional de alta energía (Kaifu, 2017).
- La relación entre ambos grupos es objeto de debate. Los investigadores coinciden en que las muestras craneales son morfológicamente diferentes, pero existe controversia sobre lo que refleja esta diferencia (Baab 2010; Baab y Zaim 2017; Kaifu et al , 2008 , 2015; Zeitoun et al, 2010). Algunos investigadores suponen que la variación fue el resultado de una evolución local anagenética, mientras que otros suponen la discontinuidad del linaje evolutivo y, por lo tanto, el reemplazo de la población de H. erectus por Homo soloensis (Kaifu, 2017).
 |
Evolución de características craneales peculiares en la serie cronológica del Homo erectus de Java .
Izquierda, Sangiran 17 (0,8 Ma).
Centro, Sambungmacan 4 (~ 0.27 Ma).
Derecha, Ngandong 12 (~ 0.1 Ma?).
Sambungmacan 4 exhibe un toro supraorbital engrosado lateralmente y parietales redondeados (indicados por las puntas de flecha), dos de los muchos rasgos característicos de Ngandong, mientras que conserva una morfología primitiva de la bóveda craneal baja similar a la de Sangiran/Trinil.
|
 |
| Ngandong 1 |
 |
Este de Java.
1 Sangiran
2 Sambungmacan
3 Sonde
4 Trinil
5 Ngandong
7 Kedung Brubus
8 Kalibeng
9 Kabuh
10 Pucangan
11 Mojokerto (Jetis-Perning)
|
Por Adam van Arsdale
Bibliografía:
- Jacob, Teuku. "Palaeoanthropological discoveries in Indonesia with special reference to the finds of the last two decades." Journal of human evolution 2.6 (1973): 473-485.
- Holloway, Ralph L. "Indonesian “Solo”(Ngandong) endocranial reconstructions: Some preliminary observations and comparisons with Neandertal and Homo erectus groups." American Journal of Physical Anthropology 53.2 (1980): 285-295.
- Swisher, Carl C., et al. "Age of the earliest known hominids in Java, Indonesia."Science 263.5150 (1994): 1118-1121.
- Swisher, C. C., et al. "Latest Homo erectus of Java: potential contemporaneity with Homo sapiens in southeast Asia." Science 274.5294 (1996): 1870-1874.
- Anton, Susan C. "Endocranial hyperostosis in Sangiran 2, Gibraltar 1, and Shanidar 5." American journal of physical anthropology 102.1 (1997): 111-122.
- Antón, S. C. "Cranial evolution in Asian Homo erectus: the Ngandong hominids."Geological Research Development Centre Special Publication 27 (2001): 39-46.
- Kaifu, Yousuke, Fachroel Aziz, and Hisao Baba. "Hominid mandibular remains from Sangiran: 1952–1986 collection." American journal of physical anthropology 128.3 (2005): 497-519.
- Dennell, R. W. "The Solo (Ngandong) Homo erectus assemblage: a taphonomic assessment." Archaeology in Oceania 40.3 (2005): 81-90.
Te puede interesar:
|
No hay comentarios:
Publicar un comentario