Las excavaciones en la cámara central de Denisova, en los Montes Altai, han revelado ocupaciones del Pleistoceno entre 125-30 ka años, con restos líticos musterienses y levalloisienses. Una datación de los huesos de fauna de la parte inferior de la capa 11, realizado en 2015 por Katherina Douka et al ha proporcionado unas fechas de 48-50 ka.
El fragmento de falange Denisova 3 fue examinado por Johannes Krause, Svante Päabo y colegas del Proyecto Genoma Neandertal en la Instituto Max Planck de Antropología Evolutiva. El ADNmt es diferente al de los seres humanos anatómicamente modernos, y al de los neandertales. A falta de otra denominación, se ha convenido tácitamente que perteneció a un "denisovano".
El ADN de varios individuos cuyos restos fueron recuperados en la cueva Denisova permite conocer que la cueva fue habitada por denisovanos entre hace 287-55 ka y por neandertales entre hace 193-97 ka (Jacobs et al, 2019). Los estudios paleogenéticos han revelado que el ADN denisovano se puede encontrar en algunas poblaciones vivas, particularmente en Melanesia y el norte de Australia y han calculado las fechas de divergencia con los neandertales y los humanos modernos y de diversos eventos de hibridación. Denisovanos y neandertales divergieron de un antepasado común hace 390 ka. Entre ambos grupos hubo episodios de hibridación. Asimismo, los denisovanos se mezclaron con los antepasados de los actuales humanos de Oceanía y Asia. Por otra parte, los denisovanos recibieron el aporte genético de uno Homo arcaico que divergió hace más de 1 Ma de los antepasados de los humanos actuales.
Se han recuperado muy pocos restos de Homo, entre ellos (dataciones de las capas estratigráficas de Jacobs et al, 2019),
Denisova 2. Segundo molar inferior deciduo muy desgastado recuperado en 1984 en la capa 22.1 de la Galería Principal, datada en más de 287 ka. Según el análisis genético (ADNmt y ADN nuclear), perteneció a una niña denisovana que vivió entre hace 139-184 ka (Slon et al, 2017). La morfología indica una edad de 10-12 años.
Denisova 3. En 2008, un equipo dirigido por los arqueólogos Michael Shunkov y Anatoli Derevianko de la Academia Rusa de Ciencias de Novosibirsk encontró en la capa 11.2 de la cámara Este, una banda gruesa de piedra arenisca, una fragmento distal de falange de un infantil. La capa se ha datado en más de 55 ka, mientras que la datación por medios genéticos es de hace ca. 85 ka.
En 2010, el equipo de Max Planck secuenció el 70% del genoma nuclear. El genoma quedó completado en 2012.
Según los análisis genéticos, la falange perteneció a una joven denisovana de pelo y ojos castaños y tez oscura. La población era reducida y muy endogámica.
El genoma incluye trazas de un homínido desconocido que divergió hace 4-1 Ma del ancestro de denisovanos, neandertales y HAM.
Según estos estudios, completados por el equipo de David Reich (2011), el ADN de los denisovianos comparte ciertas mutaciones con el de los melanesios de Papúa Nueva Guinea y la isla de Bougainville, los aborígenes australianos, los nativos de Fiji, los indonesios del este, y los Mamanwa (Filipinas) que no se encuentran en los neandertales u otras poblaciones modernas. Tampoco se encuentra en los asiáticos del este del continente, los indonesios occidentales, los Jehai (Malasia), y los Onge (Islas Andamán). Estos resultados sugieren que los denisovanos vivieron dentro de una extraordinariamente amplia gama geográfica y ecológica, desde Siberia hasta el Asia tropical y que los ancestros de los asiáticos orientales no estaban en el sudeste de Asia, cuando se produjo el flujo de genes denisovanos.
Denisova 4. Créditos: Bence Viola.
Denisova 4. Molar de macho adulto hallado en la capa 11.1 de la cámara Sur (menos de 47 ka). Un análisis del ADNmt confirmó su adscripcion denisovana (Viviane Slon et al, 2015). El ADN nuclear fue secuenciado por Sawyer et al, 2015.
Denisova 5. Falange del pie hallada en la capa 11.4 de la cámara Este (más de 105 ka) que perteneció a un neandertal, en base al análisis genético. La datación por medios genéticos es de hace ca 123 ka.
Denisova 8. Probablemente un M3 superior de macho adulto, hallado en 2010 entre las capas 11.4 y 12.1 (120-105 ka). Es similar a Denisova 4 en su gran tamaño y en el mosaico de rasgos de HAM y neandertales que presenta. El ADN nuclear y mitocondrial fue secuenciado por Sawyer et al, 2015 y corresponde a un denisovano.
Denisova 9. Capa 12.2 de la cámara Este. 141-129 ka. Neandertal.
Denisova 11. Fragmento de hueso largo de una mujer de al menos 13 años de edad, hallado en la capa 12.3 de la cámara Este, datado en menos de 141 ka. El ADN-mt corresponde a un neandertal (Brown et al, 2016), pero el ADN nuclear ha revelado que el padre del individuo era denisovano, con algún ancestro neandertal. La madre procedía de un población neandertal diferente a la del ancestros del padre, más cercana a neandertales europeos posteriores que a los neandertales más antiguos procedentes de la misma cueva. (Slon et al, 2018). La datación genética es de hace más de 90 ka.
Denisova 13. Se ha anunciado el hallazgo en 2016 de un fragmento de parietal con ADN-mt denisovano en la capa 22, datada en hace 280-250 ka.
En la campaña de 2020, se recuperaron un fragmento de hueso en la capa 13 y otro en la 14 (hace 120-130 ka). En la capa 22, un molar en la parte superior (170-190 ka) y un diente deciduo en la parte inferior (250 ka).
Estratigrafía y dataciones de Denisova. Los conjuntos de artefactos están indicados a la derecha de cada secuencia. Las condiciones climáticas se han inferido de los registros de polen y fauna. Dataciones estimadas según un modelo bayesiano con el 95,4% de probabilidad. Jacobs et al, 2019.
Slon et al (2017) han conseguido detectar ADNmt denisovano y neandertal en las capas más profundas de la Cueva Denisova, en concreto de neandertales en la capa 14 y de denisovanos en la capa 15.
Comparación de Denisova 4 y 8 con M3 superior de neandertal y HAM. Sawyer et al, 2015.
Denisova 8. Sawyer et al, 2015.
En la galería este, aparecieron varios objetos ornamentales, entre ellos un anillo de mármol, un brazalete de piedra verde, una diadema de marfil de mamut lanudo y dientes perforados, agujas y colmillos de mamut trabajados, de autoría dudosa. En 2018 restos de ocre y en 2019 la capa 11 (40-45 ka, con restos denisovanos) proporcionó un fragmento de figurilla de león en marfil alóctono y posiblemente pintada con ocre rojo, que es candidata a la representación zoomorfa más antigua.
Cronología de fosiles asiáticos de Homo arcaico. Kaifu, 2017.
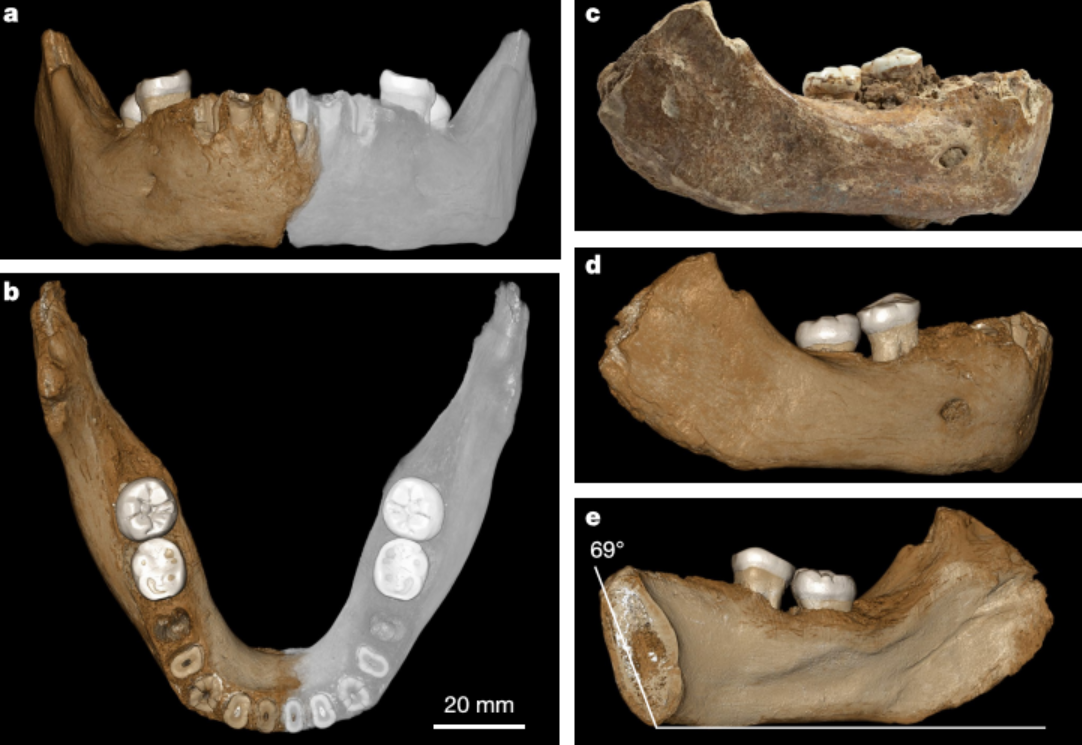
En 2019, Chen et al presentaron el hallazgo de una mandíbula, identificada como denisovana por análisis de proteínas antiguas, en la cueva de Karst Baishiya, Xiahe, Gansu, meseta tibetana, China, datada en hace al menos 160 ka mediante series U de una matriz de carbonato adherida. Por consiguiente, los denisovanos se adaptaron con éxito a ambientes hipóxicos de gran altitud mucho antes de la llegada a la región del HAM.
La mandíbula presenta un segundo molar inferior de tres raíces. Esta característica es rara en el sapiens no asiático, pero en poblaciones de origen asiático puede superar el 40% de ocurrencia. Existe pues la posibilidad de que la mayor frecuencia de la presencia de este rasgo en los asiáticos se deba a una introgresión denisovana (Bailey et al, 2019).
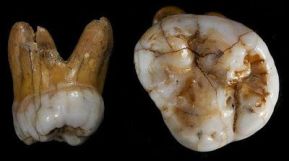
Según María Martinón-Torres et al (2017), los dientes denisovanos se caracterizan por su tamaño notablemente grande y sus rasgos morfológicos primitivos (Krause et al, 2010; Sawyer et al, 2015). A pesar de la estrecha relación filogenética entre los denisovanos y los neandertales, los molares denisovanos no muestran ningún rasgo neandertal típico. Estos dientes caen dentro del rango de variación de los fósiles del Pleistoceno Temprano y Medio del este de Asia en las cúspides mesiales bucolingualmente expandidas, la esquina distovestibular decreciente y las raíces divergentes y gruesas. Esta morfología podría interpretarse como una supervivencia tardía de un patrón típicamente asiático o como resultado de un mestizaje genético con una especie desconocida (tal vez un H. erectus superviviente ). Además, el tamaño dental está dentro del rango de los homininos del Plioceno y del Pleistoceno Temprano, lo que podría evidenciar un caso de heterosis, una consecuencia fenotípica de la hibridación genética (Ackermann, 2010; Ackermann, Rogers y Cheverud, 2006).
Otras muestras del registro fósil asiático podrían pertenecer a denisovanos, en particular Xujiayao, Penghu 1, Maba y Xuchang (María Martinón-Torres et al, 2017).
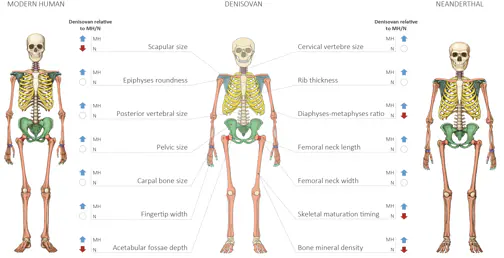
Comparación del esqueleto de HAM, denisovano y neandertal. Gokhman et al (2019).
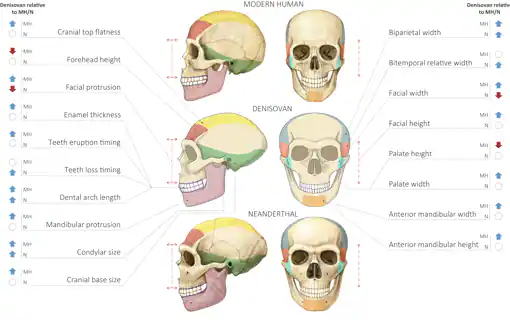
Comparación de los cráneos de HAM, denisovano y neandertal. Gokhman et al (2019).
En ausencia de fósiles que permitan especular sobre la morfología denisovana, Gokhman et al (2019) han utilizado los patrones de metilación del ADN. De acuerdo con sus resultados, los denisovanos compartían con los neandertales una cara alargada y una pelvis ancha. Como rasgos distintivos, los autores señalan un aumento del arco dental y la expansión craneal lateral. El cráneo de Xuchang, de difícil clasificación, muestra estos rasgos.
Objetos ornamentales hallados en la Cueva Denisova. Institute of Archaeology and Ethnography SB RAS.
Esto complica el paradigma según el cual Homo sapiens tiene un único origen común, ya que según lo que he entendido de tu post. Si en poblaciones acuales del Oceanía y del este de Asia sudoriental se encuentran genes denisovianos, y en el resto de poblaciones asiáticas y del resto del mundo no se encuentran, se tendrá que replantear el paradigma. Por otro lado, hay algo que no me ha quedado muy claro, en la primera parte del post dices que divergieron de los neandertales de un antepasado común hace 1 m.a., y en la segunda parte pone que se separaron hace unos 0,64 m.a. Si son 640 mil años, el antepasado bien podría ser un heidelbergensis, pero creo que esta especie no vivió por aquella zona. A 1 m.a. podría ser un erectus procedente del este asiático. Aunque he leído que por su ADNm no estaría relacionado ni con la dispersión de H. sapiens desde África ni de la primera dispersión de H. erectus. Realmente la cosa es complicada. Podría ser descendiente de una posible dispersión desde África de homininos tipo Rhodesia o Bodo (?). Francesc Ribot
Gracias, Francesc Ribot. Hace 1 Ma los denisovianos y los neandertales divergieron de los humanos modernos. Hace 640 ka se produjo la divergencia entre denisovianos y neandertales. Modifico el post para eliminar la confusión. Quizá el concepto de especie como grupo de individuos que pueden tener descendencia fértil nubla un poco las explicaciones sobre el origen del Homo sapiens. Puede que existiesen eventos esporádicos de cruzamiento genético entre los Homo de cabeza grande (erectus, heidelbergensis, neandertalensis, sapiens, etc.) que coexistieron en algún momento y lugar y con ello el taxón Homo sapiens sería inválido o bien los incluiría a todos ellos. Pero podemos utilizar otro concepto de especie, basado en la morfología y el comportamiento más que en la genética y en este caso el hecho de compartir parte del genoma no sería lo más importante. El antepasado común de denisovianos y neandertales tuvo que ser heidelbergensis, coincido contigo. Por otra parte, si sapiens se separó hace 1 Ma de neandertales y denivosianos, sapiens no desciende de heidelbergensis, también coincido.
Muchas gracias por el aporte. Interesantísimo. Hay algo que no entiendo y probablemente sea muy obvia la respuesta. Yo ya he leído esta entrada del blog anteriormente. Ahora aparece de nuevo con en portada con la fecha actualizada. Así que entiendo que debe tener contenido adicional. Pues bien ¿hay alguna forma rápida de saber cuál es exactamente ese nuevo contenido sin volver a leer el artículo e intentar deducirlo? Mis disculpas si es algo demasiado obvio.
Blogger no dispone de ninguna herramienta que permita discriminar los contenidos por su fecha. Sin embargo, hay un "truco" para localizar los contenidos más recientes, que es útil en la mayoría de los casos: introducir en el buscador del navegador el año en curso y luego el año anterior, ya que los contenidos suelen incluir las referencias o enlaces a las publicaciones científicas que los soportan. En este caso, la búsqueda de 2019 nos señalará las dataciones de Jacobs et al, el hallazgo de un fragmento de figurilla de león, la mandíbula de Baishiya con publicaciones de Chen et al y Bailey et al y las hipótesis sobre la morfología denisovana de Gokhman et al. Siento no disponer de otra alternativa. Muchas gracias por su interés.
El conocimiento es producto del pensamiento. Primero es la vida, después el pensar y en consecuencia el conocimiento. El conocimiento del pensar es el conocimiento del mismo conocimiento, es el pensamiento puro o prístino. El pensamiento acerca del conocimiento no es solamente episteme- gnosis ni filo-gniciones sino todo aquello que se refiera a la vida en manifestación de señal y aviso. La vida es un indicador de pensamiento y al contrario el pensamiento es un indicador de vida. Por eso todo ser viviente es autoconsciente de sí mismo. Todos se manifiestan en diferentes formas. Pero en un en un arrebato de “superioridad antropocéntrica” se ha condenado a las otras “especies” que entienden nuestro sistema de señales, lenguaje y comunicación. “Los humanos destacamos en lenguaje, pensamiento, inteligencia, memoria, percepción, atención y conciencia, dando emergencia a productos como el pensamiento simbólico, el arte o la religión y a una teoría de la mente muy superior a la del resto de los animales.” El cálculo y la lógica actuaron como fundamentos abstractos (perceptuales síquicos) para el desarrollo de las “matemáticas” (conocimientos de mecanismos) en sus inicios de medición, valoraciones cualitativas de las cosas y en su posterior desarrollo de las cantidades, volúmenes y formas y que permitieron a las primeras comunidades asegurar alimentos y pensar la seguridad colectiva. El pensamiento de los primeros ancestros fue lógico, abstracto y concreto y dejaron evidencias hoy indescifrables para la filosofía antropológica.
Muchas gracias a Juan Manuel Fernández López. El Blog va muy bien para profundizar cuando lees, por ejemplo, buenos libros como el tan vendido de Arsuaga. Se agradece la inclusión de libros y artículos en español e inglés. Y pediría que si el Blogger juzga como especialmente recomendable un título, por favor que lo indique.






Esto complica el paradigma según el cual Homo sapiens tiene un único origen común, ya que según lo que he entendido de tu post. Si en poblaciones acuales del Oceanía y del este de Asia sudoriental se encuentran genes denisovianos, y en el resto de poblaciones asiáticas y del resto del mundo no se encuentran, se tendrá que replantear el paradigma.
ResponderEliminarPor otro lado, hay algo que no me ha quedado muy claro, en la primera parte del post dices que divergieron de los neandertales de un antepasado común hace 1 m.a., y en la segunda parte pone que se separaron hace unos 0,64 m.a. Si son 640 mil años, el antepasado bien podría ser un heidelbergensis, pero creo que esta especie no vivió por aquella zona. A 1 m.a. podría ser un erectus procedente del este asiático. Aunque he leído que por su ADNm no estaría relacionado ni con la dispersión de H. sapiens desde África ni de la primera dispersión de H. erectus. Realmente la cosa es complicada. Podría ser descendiente de una posible dispersión desde África de homininos tipo Rhodesia o Bodo (?).
Francesc Ribot
Gracias, Francesc Ribot.
ResponderEliminarHace 1 Ma los denisovianos y los neandertales divergieron de los humanos modernos. Hace 640 ka se produjo la divergencia entre denisovianos y neandertales. Modifico el post para eliminar la confusión.
Quizá el concepto de especie como grupo de individuos que pueden tener descendencia fértil nubla un poco las explicaciones sobre el origen del Homo sapiens. Puede que existiesen eventos esporádicos de cruzamiento genético entre los Homo de cabeza grande (erectus, heidelbergensis, neandertalensis, sapiens, etc.) que coexistieron en algún momento y lugar y con ello el taxón Homo sapiens sería inválido o bien los incluiría a todos ellos. Pero podemos utilizar otro concepto de especie, basado en la morfología y el comportamiento más que en la genética y en este caso el hecho de compartir parte del genoma no sería lo más importante.
El antepasado común de denisovianos y neandertales tuvo que ser heidelbergensis, coincido contigo. Por otra parte, si sapiens se separó hace 1 Ma de neandertales y denivosianos, sapiens no desciende de heidelbergensis, también coincido.
Muchas gracias por tu clarificadora nota.
ResponderEliminarThanks.
ResponderEliminarMuy interesante...gracias.
ResponderEliminarMuchas gracias por el aporte. Interesantísimo.
ResponderEliminarHay algo que no entiendo y probablemente sea muy obvia la respuesta. Yo ya he leído esta entrada del blog anteriormente. Ahora aparece de nuevo con en portada con la fecha actualizada. Así que entiendo que debe tener contenido adicional.
Pues bien ¿hay alguna forma rápida de saber cuál es exactamente ese nuevo contenido sin volver a leer el artículo e intentar deducirlo?
Mis disculpas si es algo demasiado obvio.
Muchas gracias por su trabajo.
Blogger no dispone de ninguna herramienta que permita discriminar los contenidos por su fecha. Sin embargo, hay un "truco" para localizar los contenidos más recientes, que es útil en la mayoría de los casos: introducir en el buscador del navegador el año en curso y luego el año anterior, ya que los contenidos suelen incluir las referencias o enlaces a las publicaciones científicas que los soportan.
EliminarEn este caso, la búsqueda de 2019 nos señalará las dataciones de Jacobs et al, el hallazgo de un fragmento de figurilla de león, la mandíbula de Baishiya con publicaciones de Chen et al y Bailey et al y las hipótesis sobre la morfología denisovana de Gokhman et al.
Siento no disponer de otra alternativa.
Muchas gracias por su interés.
Muchas gracias por la respuesta y, de nuevo, por todo tu trabajo.
EliminarGracias a ti. Cordiales saludos.
EliminarEl conocimiento es producto del pensamiento. Primero es la vida, después el pensar y en consecuencia el conocimiento. El conocimiento del pensar es el conocimiento del mismo conocimiento, es el pensamiento puro o prístino.
ResponderEliminarEl pensamiento acerca del conocimiento no es solamente episteme- gnosis ni filo-gniciones sino todo aquello que se refiera a la vida en manifestación de señal y aviso. La vida es un indicador de pensamiento y al contrario el pensamiento es un indicador de vida. Por eso todo ser viviente es autoconsciente de sí mismo. Todos se manifiestan en diferentes formas.
Pero en un en un arrebato de “superioridad antropocéntrica” se ha condenado a las otras “especies” que entienden nuestro sistema de señales, lenguaje y comunicación.
“Los humanos destacamos en lenguaje, pensamiento, inteligencia, memoria, percepción, atención y conciencia, dando emergencia a productos como el pensamiento simbólico, el arte o la religión y a una teoría de la mente muy superior a la del resto de los animales.”
El cálculo y la lógica actuaron como fundamentos abstractos (perceptuales síquicos) para el desarrollo de las “matemáticas” (conocimientos de mecanismos) en sus inicios de medición, valoraciones cualitativas de las cosas y en su posterior desarrollo de las cantidades, volúmenes y formas y que permitieron a las primeras comunidades asegurar alimentos y pensar la seguridad colectiva.
El pensamiento de los primeros ancestros fue lógico, abstracto y concreto y dejaron evidencias hoy indescifrables para la filosofía antropológica.
Muchas gracias a Juan Manuel Fernández López. El Blog va muy bien para profundizar cuando lees, por ejemplo, buenos libros como el tan vendido de Arsuaga. Se agradece la inclusión de libros y artículos en español e inglés. Y pediría que si el Blogger juzga como especialmente recomendable un título, por favor que lo indique.
ResponderEliminarEste era yo, josepsolepuig20@gmail.com
ResponderEliminarMmorpg Oyunlar
ResponderEliminarINSTAGRAM TAKİPÇİ SATIN AL
Tiktok Jeton Hilesi
TİKTOK JETON HİLESİ
Sac ekimi antalya
referans kimliği nedir
instagram takipçi satın al
METİN2 PVP SERVERLER
instagram takipçi satın al
En son çıkan perde modelleri
ResponderEliminarsms onay
mobil ödeme bozdurma
nft nasıl alınır
Ankara Evden Eve Nakliyat
TRAFİK SİGORTASİ
Dedektor
Site kurmak
aşk kitapları
Really appreciated reading this type of innovative blog. BookMyEssay gives the best UK Linux Assignment Help.
ResponderEliminar