 |
| Huellas de Laetoli. National Geographic. Kenneth Garrett. |
Se duda si los primeros Hominini se desplazaban mediante una bipedación desarrollada o si, por el contrario, mostraban una bipedación parcial, un estadio intermedio, con rasgos primitivos, con persistencia de formas
de locomoción anteriores.
Dos son los rasgos morfológicos que pueden servir para dar una respuesta:
Dos son los rasgos morfológicos que pueden servir para dar una respuesta:
v
la
forma de la cadera
o
Hay motivos de orden
biomecánico que permiten pensar (Deloison) que la rotación necesaria del eje
del cuerpo al enderezarse pasa por la pelvis y, por tanto, es muy posible que
empezasen por esa parte del cuerpo las transformaciones que condujeron a la bipedación.
§
Del
examen de los restos de las caderas de los Australopithecus
de África del Este y de Sudáfrica, Poirier concluye que su pelvis contrasta
fuertemente con la de los simios y se aproxima mucho a la humana.
o
Existe
un consenso en que la evolución funcional de la cadera, y la locomoción, ha
tenido lugar a través de cambios del ilio. Así que será, sobre todo, en el ilio
donde haya que fijarse para comparar especímenes.
§
El ilio
de los grandes simios africanos es mucho más largo y estrecho que el de los
humanos. Ese ensanchamiento y acortamiento en nuestra especie es el resultado
de la adaptación al bipedismo, de la necesidad de una inserción muscular
diferente para lograr el equilibrio necesario en la postura erecta. En las
hembras, la pelvis debe permitir el desarrollo completo del feto y su
nacimiento de lo que se deriva una anatomía de la cadera diferente en las
hembras y los machos, con diseños que podrían considerarse menos forzados por
la postura bípeda en el caso de las mujeres.
o
Disponemos
de dos pelvis de Australopithecus casi
completas: AL 288-1 Lucy (A. afarensis) y STS-14 (A. africanus). El estudio realizado por
Lovejoy y otros investigadores llega a la conclusión de que la morfología de la
pelvis era ya en esos antiguos Hominini muy similar a la humana actual. Al
tener los Australopithecus una
capacidad craneal menor, el canal de alumbramiento podía ser más pequeño y por
consiguiente las pelvis de las hembras estaría menos forzada por la
contradicción entre bipedismo y parto.
§
La idea
de que la pelvis muy ancha de los australopitecos les concede ventajas
superiores a la hora de una locomoción bípeda ha sido negada por Christine
Berge (1991). Parece que la locomoción deducible de la cadera de Lucy supone la
presencia de un comportamiento en parte arborícola.
§
Kadanuumuu,
hallado en 2005, es bípedo.
 |
En humanos y Australopithecus, la superficie isquiática está inclinada cerca de su punto medio para facilitar la posición erguida y los tendones de la corva han sufrido una transformación para la bipedestación avanzada, mientras que son primitivos en el chimpancé y el Ar. ramidus.
Los tres ilios Hominini son verticalmente cortos y anchos en sentido horizontal, formando una escotadura ciática mayor que está ausente en Pan.
Un punto de crecimiento adicional en la espina ilíaca antero-inferior también está ausente en Pan.
De C. Owen Lovejoy. 2009.
|
v
la morfología de las extremidades inferiores. Los miembros posteriores también sirven
para indicarnos si estamos ante un ser bípedo o cuadrúpedo. Se ha intentado vincular la mofología de la mano con la del pie, basándose en una supuesta influencia de la bipedación en el uso de herramientas o por el contrario en base a una bipedación causada por la necesidad de fabricar y usar herramientas. Sin embargo, parece que la bipedación y la fabricación y uso de herramientas evolucionaron independientemente (Teruo Hashimoto et al 2013). El
conjunto de la tibia y el peroné, por un lado, y los huecos del pie, por otro,
proporcionan ciertos indicios esenciales de cara a entender la biomecánica de
la locomoción. El que sus articulaciones permitan una movilidad muy grande es
algo que indica una estructura de los miembros inferiores apta para trepar. La
bipedia exige, por el contrario, articulaciones más sólidas y capaces de soportar
la distribución del peso en la postura erecta.
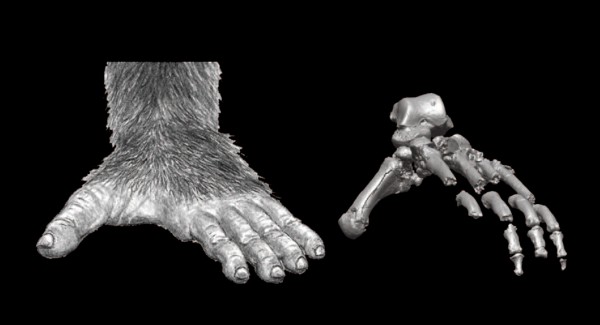 |
| Pie de Ardipithecus ramidus. C. Owen Lovejoy. 2009. |
o
En 1978
May Leakey y su equipo descubrieron en Laetoli las huellas dejadas por unos
seres bípedos hace 3,6 Ma en una capa de cenizas expulsadas por el volcán
Sadiman, humedecida por la lluvia y endurecida al secar al sol. Nuevas cenizas
enterraron y preservaron las huellas. En otros niveles de Laetoli se
encontraron restos de Australopithecus
afarensis, por lo que se cree que las huellas corresponden a individuos de
esta especie. La mayor diferencia entre la huella que deja el pie de un bípedo
y la de un cuadrúpedo estriba tanto en el arco longitudinal, muy marcado en el
humano, como en el dedo gordo, que se separa en sentido oblicuo en el
chimpancé. Muchos investigadores aceptan estas huellas como pertenecientes a
seres bípedos. Por el contrario, Yvette Deloison extrae la conclusión de que la
huella de Laetoli muestra una forma intermedia entre chimpancé y humano. En
cuanto al contorno, más parecida al chimpancé. Existen rasgos que permiten
suponer la funcionalidad prensil del pie. En 2011 un equipo dirigido por Robin
Crompton, utilizando la tecnología láser, comprobó que aquéllos individuos
caminaban de forma totalmente erguida, impulsándose con la parte delantera del
pie, como los humanos modernos. En 2011 Matthews, Breithaupt y Charles Musiba
determinaron que había un conjunto de huellas de tres individuos y otro de un
individuo que cargaba el peso en el pie izquierdo.
 |
| Huella de Laetoli |
o
Little foot está formado por un astrágalo, escafoides,
cuneiforme y parte de un metatarsiano. Procedente de Sterkfontein (Sudáfrica,
3,5-3 Ma) permaneció en una caja con otros huesos de cercopitecos durante
quince años hasta que fue redescubierto por Ronald J. Clarke en 1994 y
catalogado como Stw 573. Después, aparecieron hasta doce piezas más. Rastreando
en la gruta de procedencia se identificó embebido en la piedra lo que parece
ser el esqueleto más completo de un australopitecino. La morfología de Little Foot muestra una mezcla de rasgos
simiescos y humanos. El pie sería, por lo que hace al talón, el de un bípedo
que soporta el peso del cuerpo sobre sus pies. Pero la bipedia no estaría del
todo desarrollada: Little Foot
retiene en la parte delantera del pie la capacidad prensil necesaria para una
vida arbórea. Para Tobias (1997) eso supone tener que aceptar que mientras el
astrágalo había tomado el camino de la remodelación propia de la bipedia
habitual, parte del escafoides, el primer cuneiforme y la base del primer
metatarsiano no habían hecho lo propio y retenían aún rasgos simiescos.
Estamos, pues, ante un estadio intermedio correspondiente a un sistema
ecológico de bosque, con mosaicos de pradera intercalados, en el se produce la
conversión evolutiva del pie adaptado a la vida arbórea, con un dedo gordo
divergente y prensil, en una extremidad adecuada para la bipedia habitual. Otros
paleoantropólogos consideran que la posición de dedo gordo del pie es muy
similar a la de los humanos modernos.
 |
| Little Foot |
o
Por lo
que hace a África del Este, el yacimiento de Hadar (Etiopía) proporciona
especimenes que indican cuál es la morfología del pie de los Australopithecus
afarensis. En la serie 333 en donde se encontró a Lucy, se detectan rasgos
intermedios que colocan la morfología del pie de Australopithecus afarensis entre la humana y la de los chimpancés.
Tiene una antigüedad de entre 3,22-3,18 mda.
o
Un
cuarto metatarso del pie izquierdo de afarensis
en Hadar (AL 333-160 datado en 3,2 mda), informado en 2011 por Carol Ward,
de la Universidad de Missouri, y otros colegas. La rigidez del arco del pie, es
similar a la de los humanos modernos, en contraposición a los pies de los
simios que son más flexibles para asirse a las ramas de los árboles
 |
| AL-333-160 |
 |
| Relaciones angulares en el metatarso. |
o
OH8 de
Olduvai (Tanzania) ha sido clasificado como Homo
habilis y datado en 1,85-1,75 Ma (Patel el at, 2018). Deloison considera el pie funcionalmente
moderno.
o
La
proliferación de estudios que muestran la presencia de rasgos simiescos en el
aparato locomotor de los Hominini más antiguos puede resultar una vía muerta
para explicar la conducta de los primeros miembros de nuestra familia si se
considera que, epistemológicamente hablando, la presencia de grados intermedios
no tiene sentido evolutivo: la persistencia de rasgos primitivos, no es
significativa. Existe un argumento fuerte a favor de la gradualidad del proceso
adaptativo que condujo al bipedación, y es la presencia de los rasgos
morfológicos intermedios entre los humanos y los chimpancés en tres seres tan
separados entre sí como son los A.
Afarensis de la serie 333 de hadar, Little
Foot de Sterkfontein y OH 8 de Olduvai. Los rasgos simiescos son más
acusados en los ejemplares muy antiguos que en el de Olduvai. Pero eso es
precisamente lo que cabría esperar de un proceso de adquisición gradual de la bipedación.
Parece dudoso, pues, que la presencia de etapas intermedias en una bipedia
adquirida poco a poco vulnere los principios metodológicos de la teoría del
vector direccional.
§
Susman
y Stern aventuraron cómo fue progresando la bipedia:
·
Australopithecus afarensis y A.
africanus: conducta parcial arbórea muy clara.
·
Paranthropus robustus: Bípedo completo que ha perdido la
capacidad de trepar con agilidad.
·
Homo habilis: Bípedo por lo que hace a los pies, pero
con unos dedos de las manos funcionalmente aptos para la trepa.
o
Kadanuumuu
(Australopithecus afarensis), datado
en 3,58 mda., era totalmente bípedo.
No hay comentarios:
Publicar un comentario