Hace 40 ka, debido a la retención de agua en los glaciares,
Nueva Guinea, Australia y Tasmania formaban un único continente conocido como
Sahul. La distancia a Asia era de unos 100 km. Los primeros emigrantes debían
conocer la construcción de balsas, pues llegaron también a Nueva Bretaña y
Nueva Irlanda en el mismo periodo, separadas también a 100 km. Otra posible ruta pasa por Nueva Guinea (Bird et al, 2019). En el sureste
de Asia, las señales de humanos modernos tempranos son escasas.
- En la isla de Flores tenemos material arqueológico en Liang Bua datado entre 74-61 ka, que seguramente puedan encajar a Homo sapiens, auque hay que recordar que en esta zona es donde aparecieron los famosos restos del Homo floresiensis pero con dataciones posteriores.
- Una datación reciente de unos dientes hallados por Dubois en Lida Ajer (Padang, costa oeste de Sumatra) los ha fechado en hace 73-63 ka (Westaway et al, 2017).
- En Tam Pà Ling (Cueva de los Monos), Montañas Annamite (norte de Laos), se han recuperado (Demeter et al, 2017):
- Datados en hace 63-44 ka.
- TPL 1. Un cráneo recuperado en 2009 con morfología completamente moderna. El escaso desgaste de los molares sugiere que perteneció a un adulto joven.
- TPL 2. Una mandíbula hallada en 2010, con rasgos en mosaico.
- TPL 3. Mandíbula. Datada en hace 70 ka.
- TPL4. Fragmento de costilla izquierda.
- TPL5. Fragmento de falange del hállux derecho.
- No se han hallado artefactos.
- La morfología mosaico de TPL 2, junto con la morfología completamente moderna de TPL1 y las características de los fósiles de otros yacimientos, sugieren una gran variación morfológica en las primeras poblaciones humanas modernas que residían en Eurasia Oriental en MIS 3 (Fabrice Demeter et al, 2015).
- En la capa 14 de la Cueva Callao (norte de Luzón, Filipinas) se halló, en 2007, CCH1, un metatarsiano humano. En la misma capa estratigráfica se recuperaron posteriormente siete dientes maxilares postcaninos (CCH6-a a CCH6-e, CCH8 y CCH9), dos falanges de la mano (CCH2 y CCH5), dos falanges del pie (CCH3 y CCH4) y un eje femoral (CCH7). CCH1 y CCH6-a se dataron directamente por series de uranio a un mínimo de 67 ka y 50 ka respectivamente. Los restos pertenecen al menos a tres individuos. En base a un particular mosaico de características primitivas y derivadas, Détroit et al (2019) los asignaron a una nueva especie: Homo luzonensis.
- En Sulawesi (Célebes, Indonesia), se han recuperado útiles líticos en el sitio Leang Burung 2 datados en hace 50 ka (Brumm et al, 2018).
- En Timor oriental, en la cueva de Jerimalai, se encontraron infinidad de utensilios de piedra (más de 9.700) y casi 40.000 huesos de hasta 22 familias diferentes de pescados, además de fósiles de otros animales terrestres del entorno, de los que también se alimentaron los seres humanos, y ornamentos con una datación de hasta 42 ka. Algunos de los vestigios pertenecen a atunes, lo que implica que los habitantes de aquel lugar pescaban desde embarcaciones en alta mar. Hay que poner en relación estos pueblos con toda seguridad con los primeros colonizadores de Australia.
- Los restos más tempranos datados con seguridad, corresponden a la Cueva Niah y a Tianyuandong.
- En la Cueva Niah (Borneo), J. Harrison descubrió en 1958 un esqueleto datado en 45-39 ka. Barker et al, 2007. Klein, 2009. Curnoe et al, 2016.
- El esqueleto de Tianyuandong fue hallado en 2001 en una cueva a 6 km del núcleo del Complejo Zhoukoudian. Datado en hace 42-39 ka.
- En la Cueva Tabon (Filipinas) Fox descubrió en 1962 un hueso frontal y tres fragmentos mandibulares asociados a una industria lítica que fue bautizada como taboniense. Se dataron en hace 23 ka, aunque el contexto cronoestratigráfico estaba perturbado. Durante las excavaciones de 2000-2001 aparecieron 11 nuevos restos humanos. Junto con otros restos procedentes de excavaciones de los años 1960, suman al menos 483 piezas (Julien Corny et al, 2015). Dos M1 superiores tienen grandes dimensiones inusuales, fuera del rango de variación de lo asiáticos surorientales submodernos. Otros cuatro, caen dentro o casi de la variabilidad de los Negritos de Filipinas. La disparidad morfológica encontrada entre los M1 superiores de Tabon, es mayor que en otras poblaciones, lo que apoya la hipótesis de la presencia de dos morfotipos.
- En Wadjak
(Java), Dubois descubrió dos cráneos modernos.
- Los aborígenes australianos, neoguineanos y melanesios, comparten variaciones genéticas que no se encuentran en otras partes, y se ha calculado una población fundadora hace unos 50.000 años. Irina Pugach et al (2013) han calculado que la divergencia genética entre las poblaciones nativas de Australia, Nueva Guinea, y los Mamanwa (un grupo Negrito de las Filipinas) data de hace unos 36 ka. Hace 4230 años, estos grupos recibieron la aportación genética de poblaciones de la India, momento que coincide con un cambio de tecnología, formas de procesamiento de la alimientación y aparición del dingo en el registro arqueológico.
- Wallacea fue habitada hace 17 ka por una población de cabeza pequeña, de acuerdo con los hallazgos en Tron Bon Lei, isla de Alor, islas menores de la Sonda, Indonesia (Samper Carro et al, 2019).
- La colonización de la Polinesia y Micronesia tuvo lugar en los últimos 3000-3500 años gracias, probablemente, al desarrollo de las capacidades náuticas de las poblaciones del sudeste asiático y Australasia.
- 1500-1300 aC. Fiji.
- 1000 aC. Marianas.
- 600 aC. Cooks. Sociedad.
- 500 aC. Carolinas
- 300-50 aC. Marshalls.
- 100-0 aC. Marquesas.
- 400-600 Hawaii.
- 800 Rapa Nui.
- 1200 Nueva Zelanda.
- En cuanto a la procedencia de estos colonizadores, hay diversas teorías:
- Para Bellwood (1985) provenían de Taiwan y China continental, y su tarjeta de presentación es la cerámica tipo Lapita. Hablaban lenguas austronesias, se expandieron rápidamente, reemplazando a una población de cazadores recolectores (modelo en Dos Capas; Jacob, 1967) y tuvieron escaso contacto con los melanesios, de lenguas papúas.
- En contra de esta teoría, obran las siguientes evidencias: La cultura Lapita nunca e ha encontrado en el sureste de Asia ni en Taiwán. En Nueva Guinea hay poca cerámica Lapita y existe otra más antigua. La cerámica Lapita es tosca y mal cocida lo que no cabe esperar de una cultura procedente del sureste de Asia.
- En línea con esta teoría están los análisis de Robert Blust, quien dividió las lenguas austronesias en diez grupos. Los nativos no chinos de Taiwán hablan lenguas de nueve de estos grupos.
- También confirman esta teoría los análisis del ADNmt y los análisis genéticos de Sheila Mirabal et al (2013).
- Las poblaciones de Nueva Zelanda no tienen ascendencia taiwanesa. Tienen ascendencia en la China continental.
- El grupo Ami, el más grande en población en Taiwán, sólo afecta a grupos de Zelanda y Madagascar, pero no de Samoa y Tonga.
- Patrick Kirch entre otros, insiste en la fusión de elementos culturales melanesios y austronesios. El modelo Tripe I (intrusión, innovación, integración) defiende que la cultura Lapita surgió de diversos pueblos melanesios ligados por complejas redes sociales a pueblos del sureste de Asia durante miles de años. El grado de mezcla con las población fue escaso.
- Confirman esta teoría los análisis del cromosoma Y. En la tercera parte de los polinesios está presente en haplogrupo L, frecuente en el sur de China y en los ami de Taiwán.
- En contra de esta teoría está el hecho de que también son muy frecuentes los haplogrupos C y F, muy raros en Taiwán y sur de China.
- Stoneking aboga por un modelo de desplazamiento lento en el que los austronesios se desplazaron del sureste de Asia por Nueva Guinea mezclándose con la población y moviéndose hacia Polinesia. Curnoe et al (2016), un modelo de múltiples migraciones.
Para O'Connell et al (2018), los únicos restos bien datados en contextos confiables asignados a Homo sapiens son los que se recuperaron en Niah (Borneo) y Willandra
(Australia), con fechas de hace ca 40 ka. TPL 1 se data en hace 50-40 ka. La evidencia fósil de una presencia más temprana del HAM en la región es cuestionable.
TEORÍAS SOBRE EL POBLAMIENTO DE POLINESIA
Velocidad de desplazamiento
|
Mezcla de poblaciones
|
|
Bellwood
|
Rápida
|
Escasa
|
Triple I
|
Lenta
|
Escasa
|
Stoneking, Curnoe
|
Lenta
|
Grande
|
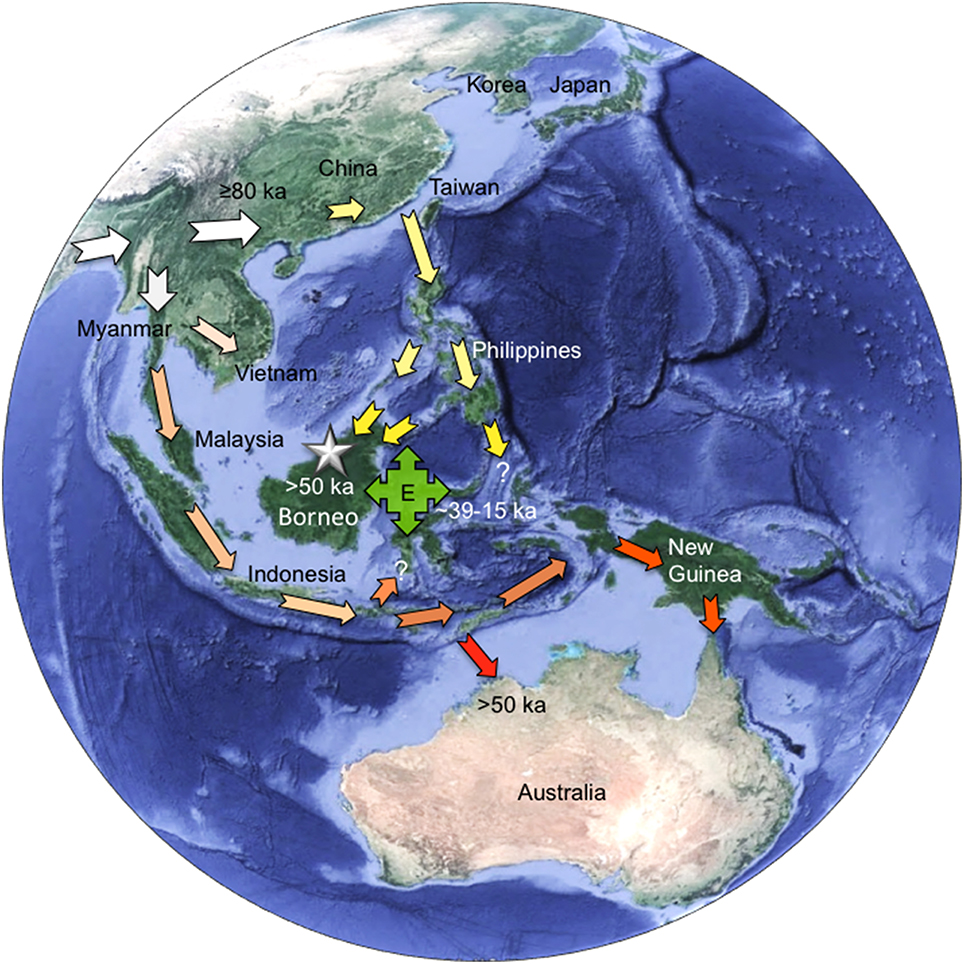 |
| Modelo de múltiples migraciones para el poblamiento del Sureste de Asia. Curnoe et al, 2016. (1) El HAM llega al sur de China hace al menos 80 ka (flechas blancas grandes). (2) Dispersiones hacia la región oceánica del sudeste de Asia a través de las Filipinas y Borneo, llegando hasta el sur de Sulawesi y Maluku (flechas amarillas). (3) Otra dispersión, a través de la península de Malasia, establece las poblaciones de los archipiélagos del sureste de Asia (flechas de color naranja claro), Australia (flechas rojas) y Nueva Guinea (flechas de color naranja oscuro). (4) El haplogrupo E aparentemente surgió hace 30 ka y se dispersó posteriormente por gran parte de la región, incluyendo Taiwán y Nueva Guinea después de hace ca 15 ka (caja verde con flechas). La estrella marca el emplazamiento de la Cueva Hiah. |
- Los ancestros comunes a euroasiáticos y aborígenes australianos divergieron de las poblaciones africanas hace 66 ka.
- La separación entre los ancestros de los australianos y los nuevaguineanos y los ancestros de los euroasiáticos sucedió hace 43 ka.
- La divergencia entre los australianos, los nuevaguineanos y el grupo Mamanwa Negrito se produjo hace 36 ka. En ese momento el Sahul constituía una sola masa y la fecha es muy anterior a la crecida de las aguas del mar que separó la isla de Nueva Guinea hace unos 8 ka.
 |
| Posibles direcciones de las migraciones humanas, localizaciones y porcentajes de mezcla. Mark Lipson et al, 2014. |
- Todos los grupos austronesios muestreados albergan ascendencia aborigen taiwanesa, más cercana que la de cualquier población continental actual.
- También han heredado ascendencia de poblaciones austro-asiáticas.
- Había una presencia sustancial austro-asiática en las islas del sudeste de Asia, antes de la llegada de los austronesios.
- La migración austronesia se produjo por el continente, mezclándose allí con los austro-asiáticos, antes de continuar hacia el oeste de Indonesia.
 |
| Yacimientos del sureste de Asia y Australia. O'Connell et al, 2018. |
 Oleadas migratorias hacia Asia Oriental, según el ADN-Y |
Australia
Las dataciones más antiguas de hace 55 ka están sujetas a controversia:
En el centro y sur del continente, disponemos de varios yacimientos con dataciones menos discutidas. En todo caso, la colonización debió de producirse mucho antes. La línea de costa del Pleistoceno estaba a cientos de kilómetros de la moderna y están inundadas extensiones considerables de tierra ocupada por los primeros colonos. Seguramente muy pocos o ningún sitio de la primera colonización han sobrevivido hasta hoy.
Hace 20 ka, en el apogeo de la última edad del hielo, los aborígenes poblaban todo el continente y habitaban la Cueva Kutikina en el río Franklin (Tasmania).
Hace 13 ka los aborígenes de Tasmania quedaron aislados del continente. Hace 8 ka Australia se aisló de Nueva Guinea y se formaron las Islas de Torres.Esta dispersión de yacimientos tempranos, se puede explicar de dos formas alternativas:
- En Moyjil (también conocido como Point Ritchie, sudoeste de Victoria) se ha descubierto una acumulación de conchas de hace menos de 125-120 ka, que podría representar un basurero de una población humana aunque no se puede descartar que fuese creada por aves marinas. Se han hallado también carbones y piedras con evidencia de fuego. Sherwood, 2018.
- Las excavaciones en Nauwalabila I y Madjedbebe (Malakunanja II) (ver Clarkson et al, 2015; Clarkson et al, 2017; O'Connell et al , 2018), al oeste de la Tierra de Arnhem, cerca de la costa norte, produjeron artefactos en sedimentos datados en hace 65-50 ka. La datación de algunos artefactos podría alcanzar los 80 ka. Investigadores como Jim O'Connell y Jim Allen (2004), advierten del movimiento vertical de los artefactos, principalmente como resultado de la actividad de las termitas.
 |
| Yacimientos del norte de Australia. Hiscock et al, 2016. |
- En 2015 se comunicó una datación de más de 50 ka en Boodie Cave, Barrow Island, Oeste de Australia, asociada a restos de alimentos consumidos por humanos. Pendiente de publicación.
- Warratyi rock shelter, southern interior. 49 ka. Evidencia más temprana en Australia de artefactos (más de hace 46 ka), ocre (49-46 ka), pigmento de yeso (40-33 ka), herramientas de hueso (40-38 ka), herramientas con mango (38-35 ka), y artefactos respaldados (30-24 ka). Hamm et al, 2016.
- La re-excavación de Karnatukul (Serpent's Glen) ha proporcionado evidencia de ocupación humana del desierto occidental australiano antes de 47,83 ka cal (Jo McDonald, 2018).
- Lake Mungo. Willandra Lakes of NSW. 42 ka. Los artefactos más antiguos se han datado por luminiscencia entre 50,1 ± 2,4 ka y 45,7 ± 2,3 ka (Bowler et al, 2003). Los restos humanos han sido fechados por datación óptica de grano simple de los sedimentos asociados en 40±2 ka (Olley et al 2006).
- Devil's Lair. Southwest Western Australia. 48 cal ka. Turney et al 2001.
- Carpenter's Gap y Riwi. Southern Kimberley. 43 cal ka. Balme 2000; Fiield et al 2001.
- Nawarla Gabarnmang, Arnhem Land, Northern Territory. 45,2 cal ka. David et al 2011.
- Parnkupirti en el centro de Australia. Veth et al 2009.
- Ganga Maya Cave, Pilbara, Western Australia. 45 ka. Pendiente de publicación.
- Bobongora, Huon Peninsula, New Guinea. 44 ka. Groube et al 1986.
- Yombon, New Britain. 40,7 cal ka. Pavlides y Gosden 1994.
- Parmerpar Meethaner. Tasmania. Steppe. 40.7 cal ka. Cosgrove 1995b.
- Ngarrabulgan. Cape York. 40 cal ka. David et al 1997.
- Puritjarra. Central Australia. 35 cal ka. Smith et al, 2001.
- Yalibirri Mindi Rock Shelter. Mid-West. Datación de hace 30 ka. de carbón asociado a herramientas líticas.
 |
| Mapa de islas del sudeste asiático que muestra las rutas de Birdsell (1977) de Sunda a Sahul. En rojo, una ruta norte con tres alternativas (1A, 1B y 1C). En azul, una ruta sur con dos alternativas (2A y 2B) también. El gris oscuro indica la extensión de las plataformas continentales hace 65 ka. Shimona Kealy, Julien Louys y Sue O'Connor (2018). |
Hace 13 ka los aborígenes de Tasmania quedaron aislados del continente. Hace 8 ka Australia se aisló de Nueva Guinea y se formaron las Islas de Torres.Esta dispersión de yacimientos tempranos, se puede explicar de dos formas alternativas:
- Una dispersión rápida por todas las regiones y paisajes de cazadores-recolectores altamente móviles.
- Una colonización más temprana. Los yacimientos corresponderían a un momento posterior, en que la actividad humana creció en importancia. Los humanos puede haber estado presentes en los paisajes por un período prolongado antes de ser visibles arqueológicamente hace unos 50 ka.
Ligado al problema de la fecha de la primera colonización y la tasa de dispersión está la discusión sobre las transformaciones ambientales causadas y en especial la reducción y extinción de la megafauna en todo el continente. Una colonización tardía implicaría que la extinción de la megafauna era casi completa en muchos paisajes antes de la aparición humana. No se han hallado lugares donde los humanos hayan dado muerte a los grandes animales extinguidos, pero esto podría ser consecuencia de la falta de conservación de los yacimientos. Las estrategias de quema de flora constituyeron la respuesta humana en ambientes fuera de control debido a la ausencia de los grandes herbívoros consumidores.
Alan N. Williams (2013), Allan N. Williams et al (2013), Tobler et al (2017) y Nagle et al (2017), han reconstruido los datos de población prehistórica de Australia.
- De la diversidad genética observada en los estudios del ADN-mt, se deduce que el grupo fundador, hace 50 ka, constó de unos mil o dos mil individuos, lo que sugiere una colonización intencionada. Los estudios genéticos indican además que no hubo episodios de colonización adicionales. Estas conclusiones son consistentes con la reconstrucción efectuada por el autor que calcula una población fundadora de mil a tres mil individuos.
- Por el contrario, Nagle et al (2015), en base al ADN-Y, defienden más de un evento a través de Nueva Guinea, para la llegada al Sahul hace ca 50 ka, así como eventos de colonización durante el Holoceno, desde la India u otros lugares.
- Nagle et al (2017) localizan el origen de los haplogrupos australianos del ADNmt quizá en la India.
- El modelo tradicional aborigen de patrilocalidad y movimiento de las mujeres a través de grandes distancias por cientos, si no miles de generaciones pueden explicar la amplia distribución geográfica de diversos haplogrupos indígenas del ADNmt (Nagle et al, 2017)
- Tras la llegada a Australia, hubo una rápida dispersión de la población que se estabilizó a niveles muy bajos.
- Entre 21-18 ka, la población se redujo hasta un 61%, seguramente como consecuencia de importantes eventos climáticos (último máximo glacial). La población correlaciona de forma estrecha con los factores climáticos y ambientales. Los modelos de asentamiento en refugios han sido confirmado por las dataciones y las técnicas geoespaciales.
- En el periodo frío de reversión del Antártico (14,5-12,5) la población se concentra nuevamente en refugios.
- Las poblaciones durante todo el Paleolítico Superior fueron bajas, con un aumento gradual lento pero sostenido desde hace 12 ka (antes de lo que se creía), coincidiendo con un aumento de nivel del mar y el fortalecimiento del monzón del norte. El crecimiento fue mayor en el sureste, gracias a una creciente utilización espacial y a la diversificación de las actividades económicas y de los lugares de residencia, con aparición de tecnología compleja.
- La población hace quinientos años era de 1,2 millones.
- En el momento del contacto con los europeos (1788), la población era de unos 770.000-1.100.000 individuos.
Anders Bergström et al (2016) han analizado el ADN-Y de una muestra de aborígenes australianos. Según el árbol filogenético de máxima verosimilitud, los papúes y aborígenes australianos (Sahul) quedaron divididos en dos clados monofiléticos C y K* (ver también Malaspinas et al, 2016; Mallick et al, 2016).
- Se infiere un tiempo de divergencia de: 54,3 ka entre K* y sus parientes más cercanos (haplogrupos R y Q) y 54,1 ka entre C y sus parientes más cercanos (haplogrupo C5). Estas fechas son consistentes con el registro arqueológico y con los estudios de todo el genoma y refutan un origen reciente procedente de India o Sri Lanka.
- Aunque no son descartables tiempos de divergencia más recientes en caso de ampliación de la muestra, la divergencia entre los aborígenes australianos y los papúes es de 50,1 ka en C y 48,4 ka en K*. No se puede precisar si esta separación sucedió después de la llegada de las poblaciones a Sahul o antes. La evidencia actual es consistente con un modelo simple de una sola entrada y posterior rápida divergencia.
- Alrededor de hace ca 6-4 ka, pequeñas herramientas de piedra comenzaron a ser utilizados ampliamente en Australia, se propagaron las lenguas Pama-Nyungan y apareció el dingo. Se ha especulado con la entrada de una población considerable en ese momento. Este estudio apoya por el contrario un origen indígena de los cambios tecnológicos y lingüísticos.
Desde el punto de vista paleontológico se ha diferenciado en Australia una población grácil y otra robusta.
- La grácil está representada por los restos de Lake Mungo. En Mungo (Nueva Gales del Sur, Australia) se recuperaron varios individuos (68-56 ka; la datación es polémica y se ha considerado alternativamente 42 ka). Los esqueletos son más esbeltos que WHL 50, con bóveda craneal más redondeada, frontal más recto, huesos de menor grosor, relieves supraorbitales poco importantes; pero también conservan rasgos robustos, principalmente en la región occipital. Esto ha servido de argumento a la hipótesis de varias oleadas de poblamiento de Australia. Se desconoce la población del sudeste asiático de la que partió el poblamiento de Australia, pues todos los restos hallados allí son robustos.
- LM1. Un esqueleto fragmentario, probablemente femenino y quemado. Es la cremación más antigua que se conoce.
- LM 3, individuo enterrado con las manos sobre el bajo vientre, los dedos entrecruzados y el cuerpo espolvoreado de ocre. El ADNmt de LM 3 es el más antiguo conocido. Es muy diferente del ADNmt de otros restos australianos (más robustos) y de los individuos actuales (curiosamente, más conectados con los restos robustos, contradiciendo lo que muestra la morfología), por lo que corresponde a un linaje que divergió. Descubierto en 1974.

Anthea Carstairs y el Dr Wilfred Shawcross descubriendo Mungo Man en 1974.
- La robusta, por los restos de Willandra y Kow Swamp. Los cráneos de esta morfología tienen algunos rasgos claramente arcaicos, como frentes aplanadas y bóvedas bajas, máxima anchura del cráneo en posición baja, toro angular en la porción posterior de las líneas temporales, toro occipital marcado, mayor prognatismo y mandíbulas y dientes de mayor tamaño.
- En Willandra (Nueva Gales del Sur, Australia) se descubrieron restos de varios individuos. WHL 50 posee un cráneo muy robusto con huesos gruesos, relieves pronunciados y frontal muy oblicuo en continuidad con el relieve supraorbital. Impresiones musculares marcadas. La anchura máxima del cráneo se sitúa abajo, inmediatamente por encima de las apófisis mastoides. 1.450 cc. No ha sido posible datarlo, pero no debe pasar de 20 ka, aunque se ha sugerido una datación de 26 ka. Otros individuos son gráciles o intermedios.
- En Kow Swamp (Nueva Gales del Sur, Australia) se hallaron los restos de 40 individuos, datados en hace 14-10 ka. Algunos cráneos fueron objeto de deformaciones artificiales. Enterramientos ricos en adornos. Uno de los individuos fue incinerado.
- Su morfología ofrece numerosas semejanzas con la de los aborígenes australianos y presenta gran variabilidad.
- Algunas características, como el grosor y robustez del cráneo, la cara grande y proyectada, los pómulos grandes, la tuberosidad en la base del pómulo, la frente plana, el borde redondeado en la base de la cuenca ocular y la falta de demarcación entre las regiones nasal e inferior, recuerdan al Homo erectus de Java.
- Otras características recuedan a WLH 50.
Algunos investigadores defienden una continuidad morfológica en la región, a partir de poblaciones de Homo erectus del sureste asiático, pero algunos aspectos morfológicos se alejan de esa posible continuidad, tales como la región occipital del cráneo (vertical y alta en sapiens, inclinada y corta en erectus), y la forma del fémur (con distintos puntos de anchura mínima entre erectus y sapiens). Westaway y Groves (2009), descartan la hibridación erectus-sapiens.
La tecnología lítica de estos pobladores es simple si se la compara con la de otros lugares de equivalente datación (ver Hiscock 2008; White, 1977). Quizá se deba al aprovechamiento de recursos marinos y de materiales orgánicos como madera, hueso y concha. Boivin et al (2013) resaltan el hecho de que Homo sapiens parece haber llegado a Australia sin el paquete africano de innovaciones y que estas las fue adquiriendo de forma independiente y poco sistemática. O'Connell y Jim Allen (2004) proponen que la simplicidad de las tecnologías australianos tempranas se correlacionan con una dieta poco amplia de alimentos con escasa manipulación y procesamiento, para los que bastaban tecnologías simples. No obstante, con dataciones cercanas o superiores a hace 40 ka han aparecido en el norte de Australia fragmentos de hachas acanaladas de borde lijado para trabajar la madera y otras actividades que seguramente se utilizaban enmangadas (Geneste et al, 2000). Además, si bien la variedad de implementos de los primeros conjuntos puede parecer limitada, por otra parte las lascas retocadas estaban a menudo relativamente estandarizadas. La existencia de instrumentos sobre hueso está documentada en yacimientos como Devil’s Lair, Bay Cave, Nunamira Cave o Bone Cave. Algunas puntas de hueso como las de Warren Cave, Tasmania, podrían haberse usado parra confeccionar prendas de vestir.
La variación cultural aumentó a lo largo de la Prehistoria, en adaptación a nuevos entornos sociales y físicos. Los modelos de poblaciones de dispersión rápida, pero de crecimiento lento defendidas por O'Connell y Jim Allen (2004) sugieren una conectividad cultural mínima entre regiones distantes durante la fase colonizadora, lo que facilitó la aparición de diferencias regionales en los sistemas culturales. El pensamiento complejo, con diversas tradiciones regionales, queda también patente en:
La variación cultural aumentó a lo largo de la Prehistoria, en adaptación a nuevos entornos sociales y físicos. Los modelos de poblaciones de dispersión rápida, pero de crecimiento lento defendidas por O'Connell y Jim Allen (2004) sugieren una conectividad cultural mínima entre regiones distantes durante la fase colonizadora, lo que facilitó la aparición de diferencias regionales en los sistemas culturales. El pensamiento complejo, con diversas tradiciones regionales, queda también patente en:
- La utilización de ocre (Lake Mungo 3 y Malakunanja II). Más común en el norte y oeste del continente, y rara vez en el sureste.
- Los collares de conchas (Mandu Mandu). Entre 45-35 ka los ornamentos, en forma de collares o pulseras de conchas o huesos perforados, teñidos con ocre, aparecen sólo en la parte noroeste del continente.
- La pintura rupestre (Laura South). Los petroglifos más antiguos se han hallado en el sur de Australia (45 ka). Los dibujos en carbón de Narwala Gabarnmung, en el Territorio Norte, están datados en 28 ka. En cuevas profundas bajo los Llanos Nullarbor en Koonalda (en el borde occidental de Australia del Sur, a unos 50 kms del mar), la población aborigen dejó hace 22 ka diseños acanalados en las paredes de la cueva.
- Las hogeras de Lake Mungo están datadas en 30,7 ka.
- La evidencia más antigua (30 ka) de cremación en todo el registro arqueológico es el cuerpo de una mujer de Lake Mungo, que fue preparado con ocre.
En Cuddy Springs antiguo lago cerca de Carinda, se han encontrado las evidencias más antiguas de panificación (30 ka). Hace 18 ka se extienden las semillas, la molienda y el horneado.
 |
| Refugios y barreras en el poblamiento de Australia |
Indonesia
Meryanne K Tumonggor et al (2013) han estudiado las variantres del cromosoma Y y el ADNmt en Indonesia.
Han encontrado evidencias de varias etapas de colonización:
- La primera etapa, forma parte de la dispersión africana hace 50 ka. El nivel del mar era entonces mucho más bajo, las islas estaban unidas en una gran masa de tierra y el occidente de Indonesa unido al continente.
- En la segunda etapa se produjeron, a lo largo del Paleolítico, episodios recurrentes de colonización desde el continente asiático.
- La tercera etapa representa los movimientos del Neolítico de poblaciones endémicas. Algunos pueden implicar dispersiones de población desde (y quizás a) Taiwan, mientras que otros reflejan los movimientos entre grupos de islas de Indonesia.
- La cuarta etapa refleja los movimientos históricos en Indonesia, en su mayoría relacionados con el comercio y la consiguiente expansión de las grandes religiones de la India, Arabia y China.
Un artículo fascinante que ilustra detalladamente acerca del caminar del ser humano sobre regiones del planeta Tierra. Con datos concretos, acompañados de Imágenes, dibujos, fotos, cuadros, etc. que actúan como soporte para la comprensión de todo lo aquí informado. Muy agradecida por haberme permitido ampliar mi cultura genera.
ResponderEliminarMuchas gracias.
EliminarEXCELENTE trabajo.
ResponderEliminar