 |
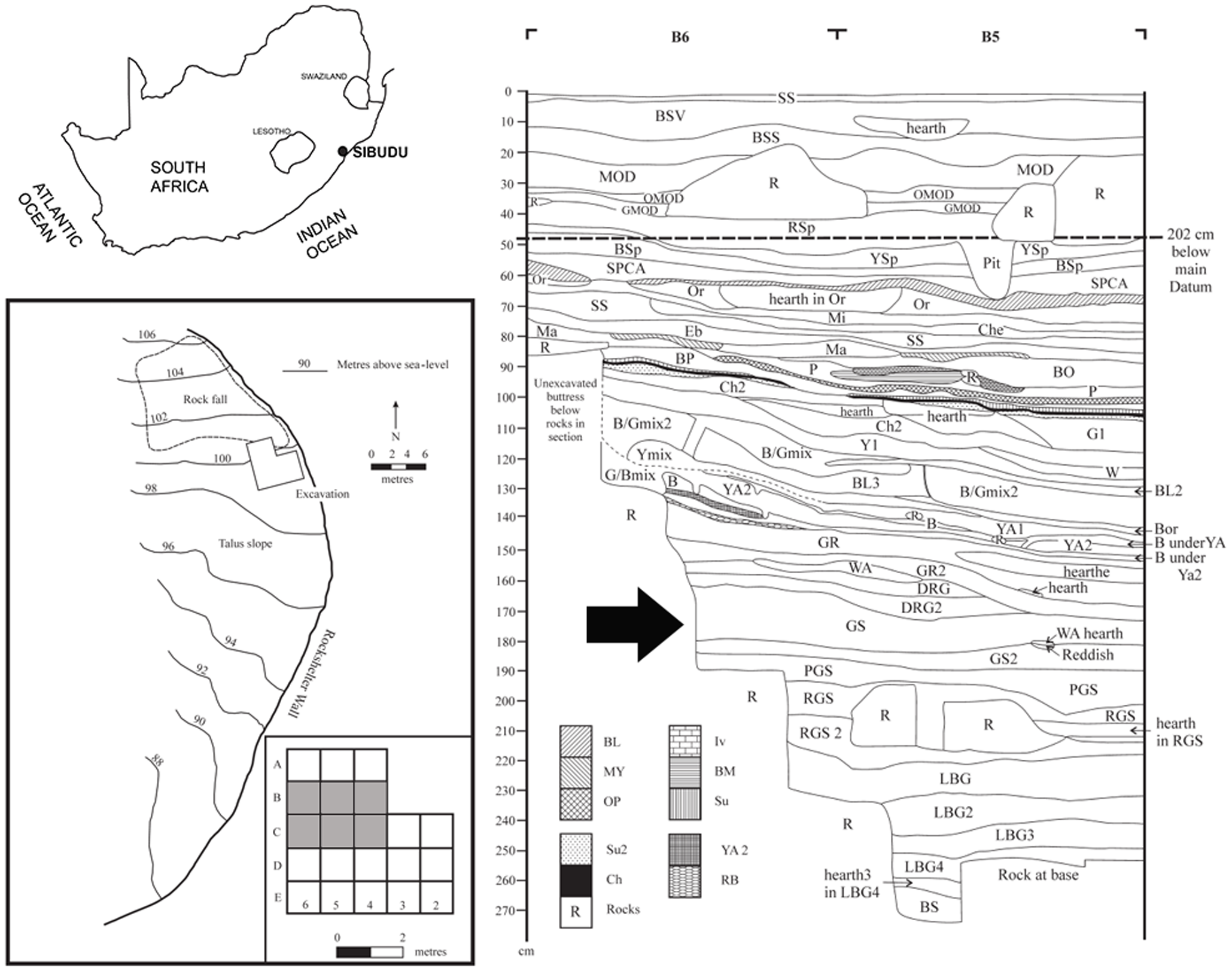
| Situación de Sibudu, retícula de excavación y estratigrafía. |
Los tecnocomplejos Howiesons Poort y Still Bay (MIS 4), se reconocen en varias zonas ecológicas de Sudáfrica. Contienen herramientas relativamente más retocadas, con cara pasiva y puntas bifaciales y muestran niveles elevados de prácticas innovadoras: tratamiento térmico de la piedra para la producción de puntas bifaciales, percusión con martillo blando, pequeños artefactos con cara pasiva, herramientas de hueso, cuentas, ocre, grabados en ocre y cáscaras de huevo de avestruz y herramientas compuestas con el uso de adhesivos incluyendo la más antigua conocida tecnología de arco y flecha. Estos niveles de innovación y complejidad se cree implican un comportamiento simbólicamente mediado (Henshilwood Marean 2003). Esta cualidad cognitiva (Wadley, Hodgkiss, y Grant 2009), pudo conferir ventaja evolutiva a los grupos de Homo sapiens que emigraron de África después de hace unos ~60 ka (Mellars 2006; Mourre, Villa y Henshilwood 2010). A lo largo del tiempo, se producen cambios sutiles en los tipos de artefactos con cara pasiva, herramientas de retoque y patrones de explotación de materias primas. Desde hace algún tiempo, la Howiesons Poort se ha considerado como una industria de transición a la LSA. La talla bipolar y las estrategias microlíticas han sido algunos de los principales rasgos utilizados para distinguir las tecnologías LSA.
El sitio arqueológico de Sibudu es un gran refugio de piedra situada sobre el río Tongati (también escrito "uThongathi") en KwaZulu-Natal a unos 40 km al norte de Durban, a 15 km del Océano Índico. Excavado por Lyn Wadley entre 1998-2011. Las excavaciones actuales están dirigidas por Nicholas Conard. El sitio conserva los depósitos que van desde la pre-SB (> 75 ka) a la MSA final (~ 37 ka): pre-Still Bay, Still Bay, Howiesons Poort, post-Howiesons Poort, MSA tardío, MSA final y Edad de Hierro. El sitio fue fechado utilizando OSL (Jacobs et al 2008).
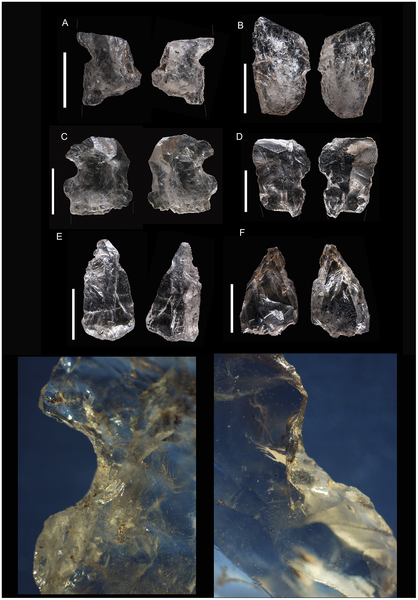 |
| Artefactos de cuarzo con muesca de la capa GS de Sibudu |
 |
| Núcleos bipolares de la capa GS de Sibudu |
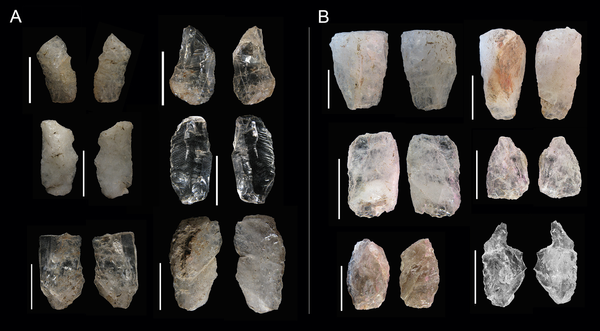 |
| Hojitas de la capa GS de Sibudu |
Paloma de la Peña y Lyn Wadley han estudiado la tecnología bifacial de cuarzo de la capa Grey Sand (GS). La gestión de este material ha resultado ser muy diferente la de otros tipos de materia prima (
corneana o dolerita). La selección de cuarzo para la de talla es notable por dos razones: en primer lugar, el cuarzo es difícil de tallar debido a sus fallas y planos de fractura (especialmente para la talla a mano alzada); en segundo lugar, el pequeño tamaño de los nódulos añade un nivel adicional de dificultad. Se ha estimado la edad de Grey Sand en 63,8 ka mediante luminiscencia estimulada ópticamente de un solo grano.
- Algunas piedras de río de cuarzo fueron talladas de forma discoidal o multifacial para la producción de positivos en escamas para puntas bifaciales.
- Los núcleos bipolares son los más comunes, seguidos de los núcleos prismáticos. Su pequeño tamaño es sorprendente. En corneana y dolerita, los núcleos bipolares son ocasionales. Se emplea la talla bipolar de forma recurrente, mientras que en corneana o dolerita su uso es ocasional. En la técnica bipolar se golpea con un gran percutor sobre el núcleo apoyado. Con ello resulta un verdadero retoque doble (uno directo y otro por contragolpe). Es muy difícil de dirigir y sus resultados no son siempre controlables. Se distingue por el doble cono de percusión que muestra el útil; uno en cada extremidad. La talla bipolar hace posible la explotación de pequeños nódulos que serían imposibles de procesar mediante talla a mano alzada y produce pequeños positivos. Su aplicación implica una economía inteligente de la materia prima.
- Se utiliza una tecnología laminar con énfasis en las pequeñas hojitas de cuarzo. En corneana y dolerita se produce en cambio una distribución bimodal de positivos pequeños y grandes.
- Las estrategias de talla parecen implicar el reciclaje sistemático y la producción deliberada de microlitos.
- Muchos de los residuos de cuarzo, y probablemente una gran cantidad de núcleos tallados a mano alzada cuando ya no era posible continuar esta técnica debido al pequeño tamaño del núcleo, se reciclaron para talla bipolar a fin de obtener pequeños positivos bipolares.
- Las herramientas en cuarzo son retocadas con mayor porcentaje que las de corneana o dolerita. Muchos positivos se utilizaron sin ningún tipo de retoque, pero algunos se transformaron en piezas bifaciales o de cara pasiva. En algunas hojitas se observan dos pequeñas muescas delicadamente retocadas en sus laterales; la función de estas herramientas es todavía un enigma.
- Los diferentes métodos de talla parecen haber sido dirigidos a diferentes conjuntos de herramientas.
Para las autoras, a pesar de que la tecnología de cuarzo Howiesons Poort de Sibudu no se ha localizado en otros contextos del HP, este conjunto no es una excepción, sino un ejemplo más de la variabilidad dentro de la MSA.
El hecho de que la talla bipolar de cuarzo sea un componente sustancial de la tecnología Howiesons Poort en la capa GS de Sibudu desmantela la noción de que esta estrategia se asocia especialmente con la LSA o se puede utilizar como un marcador para el reconocimiento de la aparición de la LSA.
Por otra parte, el descubrimiento de estas estrategias en el Howiesons Poort refuta la interpretación de una acumulación tecnológica en la MSA. La aparición y desaparición de las variantes tecnológicas deben ser vistas entonces como soluciones a problemas concretos. Por lo tanto, la talla bipolar no debe ser contemplada como un hito de "comportamiento moderno", sino simplemente como una estrategia tecno-funcional.
MSA o Modo 3















