|
Situación de la Garganta de Olduvai
|
En los últimos años, el Olduvai Paleoecology and Paleoanthropology Project (TOPPP) ha producido una amplia información de diversos yacimientos de Olduvai Bed I y Bed II y Peninj, incluyendo sitios clasificados como Olduvaienses Desarrollados y Achelenses. Quaternary International ha dedicado los
volúmenes 322-323, presentados por M. Domínguez-Rodrigo, F. Díez-Martín, A. Mabulla, E. Baquedano, HT Bunn y C. Musiba, a la reconstrucción de la paleoecología de los
Homo tempranos de Bed I durante el oduvaiense y la transición al Achelense del
erectus en Bed II.
El
erectus aparece antes de 1,7 Ma. El primer Achelense es ligeramente posterior a los primeros restos claros de
erectus (Kokiselei 4 y Konso Gardula;
Beyene et al, 2013), pero la mayoría de los restos arqueológicos hasta 1,5 Ma son Olduvaienses. Por otra parte, fuera de África,
erectus precede al Achelense durante cientos de miles de años.
Homo georgicus no contaba con tecnología achelense. De la Torre y Mora (2013) sostienen que los sitios Olduvaienses podrían atribuirse a
Homo habilis y los Achelenses a
erectus, pero la mayor parte de los sitios tanto Olduvaienses como Achelenses, comprendidos entre las tobas IIB y IIC están desprovistos de fósiles de homínidos o están asociados a restos de
H. habilis o
Paranthropus.
El Achelense es reconocido por la presencia de bifaces (Lycett y Gowlett, 2008) y en menor medida por la producción de grandes escamas (Isaac, 1969). Pero esta definición es completamente inútil para la comprensión de los conjuntos dentro de un marco ecológico. La funcionalidad de un yacimiento viene determinada por las actividades llevadas a cabo, que requieren un kit de herramientas específico. Las aproximaciones tipológicas de Leakey (1971) fueron más útiles en este sentido, porque clasificaba los sitios de acuerdo a la representación de juegos de herramientas, potencialmente indicativos de las diversas actividades.
 |
| Paleolago de Olduvai |
Según Hay (1976) En Olduvai, nueve de diez conjuntos Achelenses están lejos del paleolago y el otro es indeterminado, mientras que siete de los conjuntos Olduvaienses Desarrollados se hallaron en las orillas del paleolago y el resto son indeterminados. Esta interpretación se reproduce en Peninj donde los conjuntos achelenses se encontraron más lejos del lago que los Olduvaienses (Domínguez-Rodrigo et al, 2009).
Los hallazgos en BK y SHK muestran que los homínidos continuaron explotando animales de tamaño pequeño y mediano de forma similar a la descubierta previamente en FLK Zinj. La explotación de la megafauna, con escasa evidencia antes de 1,5 Ma, parece aumentar en Bed II.
Los problemas y hallazgos anteriormente referenciados dieron lugar a una hipótesis ecológica del origen del Achelense africano (Domínguez-Rodrigo et al, 2005; Neither Hay, 1976). Según esta hipótesis el Achelense no es un producto del erectus sino que constituye una respuesta a las oportunidades y restricciones ambientales. Las soluciones tecnológicas locales no pueden separarse de un marco contextual y regional más amplio y la tecnología a nivel de sitio es una expresión de un sistema regional que requiere información contextual para explicarse.
 |
| Reconstrucción del paisaje de Olduvai |
Los factores ambientales en Olduvai
Los indicadores paleoclimáticos muestran una tendencia a la sequía en el nivel superior de Bed I, que dio lugar a las condiciones más áridas observadas en Bed II
- Emily J. Beverly, Gail M. Ashley y Steven G. Driese han identificado una hidrología fluctuante entre 1,81-1,79 Ma indicativa de un ciclo de precesión y consistente con los registros de la Garganta de Olduvai y de todo el este de África. En este intervalo temporal se produjeron las primeras migraciones de homínidos Out of Africa huyendo de un ambiente con escasez de agua, estacionalidad de precipitación anual y grandes variaciones en las precipitaciones debido a la precesión.
- Esta hidrología en combinación con los cambios tectónicos, actuó como factor de dibujo para los homínidos y otros animales (Gail M. Ashley, Emily J. Beverly, Nancy E. Sikes y Steven G. Driese).
- Doris Barboni ha integrado toda la información obtenida sobre la paleobotánica de Laetoli, Olduvai y Peninj. Los registros sugieren una vegetación similar a la actual. Las gramíneas de tipo C4 aparecen hace ~3,7 Ma y llegan a ser dominantes poco después (~3,66 Ma), probablemente en respuesta a la disminución de las precipitaciones. Hace ~2,6 Ma los pastos xerófilos C4 documentan una fuerte aridez durante el período de intensificación de la glaciación del hemisferio norte (~2,7-2,5 Ma). Después de 2 Ma, el registro de Olduvai indica patrones de vegetación complejos de pastizales, bosques cerrados, humedales y palmares vinculados a precipitaciones oscilantes, con variación en los niveles del lago. Cuando la variabilidad del clima húmedo-seco fue extrema en África Oriental (~1,9-1,7 Ma), los manantiales de agua dulce pueden haber ofrecido un refugio para varias especies, incluidos los homínidos.
- A pesar de la competencia de los carnívoros, Charles P. Egeland muestra que otros factores como el agua, la cubierta de árboles, la tierra seca, y la disponibilidad de materias primas fueron más importantes para la selección de los lugares específicos en los que los homínidos llevaron a cabo sus actividades ya que las medidas de destrucción ósea no están relacionadas con los patrones de descarte lítico y carnicería.
 |
| Localización de Olduvai y localidades designadas por Leakey (1971) |
El registro de Olduvai
El registro de Bed II es un ejemplo particularmente significativo que sugiere que si queremos construir inferencias sólidas y significativas sobre las estrategias de vida de los homínidos, no podemos separar la tecnología de otros aspectos económicos, ecológicos, funcionales y regionales. Así, Alia N. Gurtov y Metin I. Eren han comprobado que los resultados experimentales son consistentes con la idea generalizada de que los homínidos del Paleolítico Inferior reconocían las diferencias en las propiedades físicas de los diferentes tipos de materia prima. En Olduvai, la reducción bipolar, que requiere poca o ninguna habilidad, se utiliza solo en el cuarzo y no sobre el basalto porque en aquel material el resultado es ventajoso.
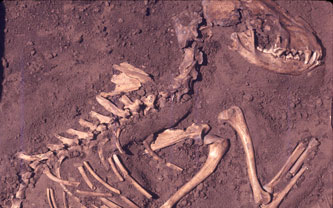
- FLK Zinj (1,8 Ma).
- Manuel Domínguez-Rodrigo, Henry T. Bunn y José Yravedra han analizado conjuntamente las marcas de corte, las marcas de dientes y las marcas de percusión, en los restos de animales de FLK Zinj y las han comparado con las obtenidas de forma experimental. El acceso primario y secundario a los cadáveres produce diferentes combinaciones de frecuencia de todas estas marcas, que pueden ser de diagnóstico del tipo de acceso. Los resultados proporcionan apoyo robusto para el acceso primario de los homínidos de FLK Zinj.
- Agness O. Gidna, Bernard Kisui, Audax Mabulla, Charles Musiba y Manuel Domínguez-Rodrigo contribuyen con un estudio longitudinal de la explotación del canal por felinos y humanos en el Parque Nacional de Tarangire.
- Henry T. Bunn y Alia N. Gurtov, proporcionan una prueba más convincente de que la caza fue la principal estrategia utilizada para obtener los cadáveres de animales: El perfil de mortalidad del conjunto está dominado por los adultos de grandes bóvidos. Además de la caza de homínidos, otros tres métodos de recolección de residuos podrían haber proporcionado estos resultados: acceso primario de carroñeros a víctimas de accidentes no relacionados con depredadores; acceso secundario de carroñeros a víctimas de depredadores; acceso primario de carroñeros agresivos a víctimas de depredadores. El perfil que domina en FLK Zinj es significativamente diferente de los perfiles de mortalidad formados por los tres métodos de barrido, lo que indica que son resultado de la caza por el Homo temprano.
- Lucía Cobo-Sánchez, Julia Aramendi y Manuel Domínguez-Rodrigo muestran evidencia convincente de que la acción lacustre o atmosférica por sí sola es insuficiente para causar la aloctonía mostrada por los conjuntos de Bed I.
- Según el estudio geomorfológico, sedimentológico, estratigráfico y geométrico efectuado por D. Uribelarrea, M. Domínguez-Rodrigo, A. Pérez-González, J. Vegas Salamanca, E. Baquedano, A. Mabulla, C. Musiba, D. Barboni y L. Cobo-Sánchez en base a 30 zanjas excavadas alrededor de FLK Zinj, el sitio se encuentra en el borde de una plataforma elevada de hábitats boscosos, rodeada de ambientes más abiertos y herbosos situados en las partes más bajas de la planicie lacustre. En el sur existían áreas de entrada, probablemente en forma de abanicos aluviales, siguiendo una dirección norte-sur. Un estudio arqueológico de las trincheras excavadas revela un agudo contraste en fósiles y de densidad de herramientas de piedra entre FLK Zinj y el paisaje circundante lo que apoya la hipótesis de que el sitio pudo haber constituido un centro de recepción de canales, carnicería y consumo.
- FLK North (1,8 Ma).
- Gail M. Ashley, Henry T. Bunn, Jeremy S. Delaney, Doris Barboni, Manuel Domínguez-Rodrigo, Audax ZP Mabulla, Alia N. Gurtov, RoniDell Baluyot, Emily J. Beverly y Enrique Baquedano interpretan los componentes del sitio FLK North, donde ha aparecido una acumulación de fósiles, cerca de un humedal dominado por la actividad de los carnívoros. FLK North está delimitado verticalmente por las tufas Ng'eju (1,818 ± 0,006 Ma) e IF (1,803 ± 0,002 Ma), y está formado por 9 niveles que comprenden un registro relativamente continuo en un ciclo de precesión de Milankovitch (seco-húmedo-seco). A medida que el lago se desvaneció durante la parte seca del ciclo, los suministros de agua superficiales disminuyeron y los manantiales y humedales subterráneos se convirtieron en el suministro de agua dulce dominante. El análisis de los huesos y artefactos fósiles recién recuperados ha demostrado que los huesos de animales grandes son en gran medida producto de la caza y alimentación de felinos y hienas.
 |
Estratigrafía del sitio SHK-1
|
- SHK (1,78-1,34 Ma).
- Manuel Domínguez-Rodrigo, Fernando Diez Martín, José Yravedra, Rebeca Barba, Audax Mabulla, Enrique Baquedano, David Uribelarrea, Policarpo Sánchez y Metin I. Eren señalan el hallazgo de una densa concentración de herramientas de piedra y restos de fauna en SHK (1,78 a 1,34 Ma). Los análisis tafonómicos sugieren el papel de un conjunto heterogéneo de agentes en la acumulación y modificación de los restos faunísticos. A pesar de ello, hay evidencia de explotación de hipopótamos y équidos por parte de los homínidos y esto pone de relieve la posibilidad de que hace 1,5 Ma los homínidos fueron diversificando su dieta y ampliando su nicho ecológico, explotando la megafauna, probablemente de manera oportunista. Un análisis comparativo de la diversidad taxonómica entre SHK frente a otros sitios superiores de Bed II apoya la noción de que el medio ambiente era bastante abierto antes del inicio de los depósitos de Bed III. El gran número de artefactos líticos contrasta con la pequeña cantidad de evidencia para el procesamiento de canales, lo que sugiere la involucración de los homínidos en diversas actividades más allá de la carnicería.
- Fernando Diez-Martín, Policarpo Sánchez-Yustos, David Uribelarrea, Manuel Domínguez-Rodrigo, Cristina Fraile-Márquez, Rosa-Ana Obregón, Isabel Díaz-Muñoz, Audax Mabulla, Enrique Baquedano, Alfredo Pérez-González y Henry T. Bunn apoyan la tendencia actual a subsumir los sitios de Bed II dentro del tecnocomplejo Achelense debido a la concurrencia de ciertos rasgos tecnológicos.
- Fernando Diez-Martín, Policarpo Sánchez Yustos, Diana Gómez de la Rúa, José Ángel Gómez González, Luis de Luque y Rebeca Barba incluyen también en el Achelense los conjuntos de ES2-Lepolosi, Sambu Escarpment, Lake Natron Basin (Tanzania), datados en 1,5–1,4 Ma.
- Fernando Diez- Martín, Policarpo Sánchez Yustos, José Ángel Gómez González, Luis Luque, Diana Gómez de la Rúa y Manuel Domínguez-Rodrigo han analizado los resultados del trabajo de campo reciente en el sitio EN1-Noolchalai, Sambu Escarpment, Lake Natron Basin (Tanzania), datado en 1,5-1,4 Ma. El sitio arqueológico se ha visto gravemente afectado por la erosión fluvial y la redeposición. Las excavaciones de 2009-2011 han descubierto una nueva colección lítica. La interpretación tecnológica confirma que los fabricantes de herramientas en Noolchalai eran conscientes de los principios volumétricos relacionados con la producción de bifaces y cuchillos.
- TK (Thiongo Korongo; 1,35 Ma).
- Manuel Santonja, Joaquín Panera, Susana Rubio-Jara, Alfredo Pérez-González, David Uribelarrea, Manuel Domínguez-Rodrigo, Audax Z.P. Mabulla, Henry T. Bunn y Enrique Baquedano ha realizado un estudio tecnológico y paleoeconómico del conjunto lítico de 5.805 piezas recuperado en el nivel inferior (TKLF). El carácter del sitio, así como las actividades llevadas a cabo en el mismo, han sido influenciados en gran medida por las materias primas utilizadas principalmente de procedencia próxima. Se pueden identificar dos diferentes cadenas operatorias: una basada en la obtención de lascas a partir de rocas volcánicas y cuarcita y otra en la fabricación de grandes y altamente estandarizados bifaces que fueron producidos, utilizados y abandonados en el sitio.
- BK (1,34 Ma).
- M. Domínguez-Rodrigo, H.T. Bunn, A.Z.P. Mabulla, E. Baquedano, D. Uribelarrea, A. Pérez-González, A. Gidna, J. Yravedra, F. Diez-Martin, C.P. Egeland, R. Barba, M.C. Arriaza, E. Organista y M. Ansón recuerdan el descubrimiento de abundantes pruebas de explotación de megafauna. La carnicería de canales tanto grandes como medianos y pequeños señalan la importancia de la carne en el comportamiento del erectus. BBK contiene la mayor cantidad de huesos modificados por homínidos y animales descuartizados de todo el registro arqueológico del Pleistoceno Temprano. La caza era una parte integral de la conducta adaptativa del erectus, aunque en la explotación de la megafauna pudieron darse más conductas oportunistas. La organización del sitio sugiere una distribución espacial diferente a la mostrada por homínidos anteriores, tal como se documenta en sitios como FLK Zinj. Esto revela la necesidad de nuevos modelos de comportamiento para explicar la funcionalidad de campamentos centrales achelenses.
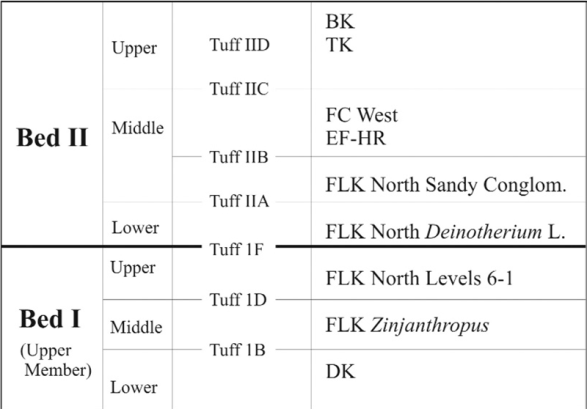 |
| Niveles de Olduvai Bed I y Bed II |
La MSA en Olduvai
El Achelense finalmente se convierte en el MSA, cuya presencia más temprana en Olduvai está documentada en Ndutu. Metin I. Eren, Adam J. Durant, Mary Prendergast y Audax Z.P. Mabulla comunican hallazgo de más de 72 lugares arqueológicos, con cientos de artefactos de la MSA recuperados. Las observaciones geológicas indican provisionalmente que Ndutu se formó por aportaciones de piroclastos y cenizas de erupciones volcánicas.

 |
| Formación de tufas en Olduvai |
Enlace
La transición olduvaiense achelense