La categoría especie es útil para la conversación trivial y para los estudios biológicos y todos entendemos lo que la palabra especie significa, pero a la hora de definirla con exactitud, aparecen dificultades derivadas del desacuerdo sobre los procesos que producen nuevas especies.
- Según el concepto tipológico una especie se define por un tipo (holotipo), básicamente morfológico (cualitativo), que se considera una referencia a la cual deben acomodarse todos los especímenes de la especie (Nelson y Platnick, 1981; Cracraft 1983; Nixon y Wheeler 1990).
- Holotipo: el espécimen u otro elemento usado por el autor o designado por él para definir una especie.
- Isotipo: un duplicado del holotipo, que forma parte de la colección original.
- Lectotipo: espécimen o elemento seleccionado a partir de material original para servir como tipo nomenclatural cuando no fue asignado un holotipo con la primera publicación o por pérdida del mismo. El lectotipo se debe elegir entre los isotipos, si no existen isotipos se debe elegir entre los sintipos, si tampoco hay sintipos se elige un neotipo.
- Sintipo: es uno de los especímenes citados originalmente por el autor que no designó holotipo o que ha enumerado simultáneamente varios ejemplares como tipos.
- Neotipo: es un espécimen o cualquier otro elemento elegido para servir de tipo nomenclatural cuando falta todo el material sobre el cual está basado el nombre del taxón.
- Paratipo: cada espécimen de una serie tipo que no sea el holotipo.
- Especies sinmórficas (sibling species): Especies prácticamente indistinguibles morfológicamente.
- Para el concepto fenético (Michener, 1970; Sokal y Crovello, 1970; Sneath y Sokal, 1973), una especie es un conjunto de organismos que se parecen entre sí y que pueden distinguirse de otros conjuntos análogos. Para caracterizar estos conjuntos se pueden utilizar métodos cuantitativos de distancia genética a partir de medidas de múltiples caracteres.
- Para el concepto biológico (Wright 1940; Mayr 1942; Dobzhansky, 1950), una especie es un grupo de individuos que se entrecruzan entre sí y que están reproductivamente aislados de otros individuos (Mayr 1942; Dobzhansky 1970).
- El flujo génico tiende a mantener las semejanzas entre los miembros de la misma especie.
- Pese a que es el más utilizado, el concepto biológico es problemático:
- Para Jerry Coyne y Allen Orr (2004) el aislamiento reproductivo no es necesariamente completo. Durante el proceso de especiación pueden producirse estados intermedios en los que el estatus de especie es más o menos irresoluble.
- No se puede aplicar de manera objetiva a las especies alopátridas (Groves, 2004).
- Es totalmente inaplicable a especies extintas (Begun y Strait, 2013).
- El concepto recognitivo de especie (especie como un sistema de reconocimiento en apareamiento) es más restrictivo que el concepto biológico e implica un sistema específico de reconocimiento de pareja o de gametos que supone la impermeabilidad del acervo genético (Paterson, 1985; Masters et al, 1987; Lambert y Spencer, 1995).
- No es aplicable a taxones en cautividad, extintos o sin reproducción sexual (Groves, 2004).
- El concepto de especie en cuanto cúmulo genético (Avise y Ball 1990; Baum y Shaw 1995; Mallet, 1995) presume que los distintos genomas se distribuyen en grupos discretos. El alelo ancestral común a la especie no se comparte con otras especies. El problema es determinar el grado de diferencia que justifica el reconocimiento de distintas especies.
- Esta definición no es válida para especies con reproducción asexual, en las que el descendiente toma sus genes de un único ascendiente, ni para los organismos procariotas, sin cromosomas.
- Para el concepto ecológico (Van Valen, 1976; Andersson 1990), una especie está delimitada por la acción de la selección natural. Una especie es un grupo de individuos que ocupan un nicho ecológico al cual están adaptados, de modo que el flujo génico se interrumpe porque la selección de nicho impide que los posibles híbridos con otros grupos análogos prosperen en los nichos parentales.
- Se dan casos de especies que intercambian sus genes, pero la selección natural se ocupa de eliminar los individuos con rasgos intermedios.
- Se dan casos de grupos de individuos aislados entre sí, es decir, que durante miles de años no han intercambiado sus genes y sin embargo son morfológicamente similares y se clasifican dentro de la misma especie.
- Para el concepto cohesivo de especie (Templeton, 1982, 1989, 1998) una especie es la población más inclusiva de individuos que tienen el potencial para la cohesión fenotípica mediante mecanismos cohesivos intrínsecos.
- Mecanismos de intercambio genético.
- Mecanismos de intercambio demográfico, que definen el nicho fundamental y los límites de la difusión de nuevas variantes genéticas por deriva genética y selección natural.
 |
| Ejemplo de homología |
- Para el concepto evolutivo de especie (Simpson 1951, 1961; Wiley, 1978; Mayden, 1997; Simpson, 1961) una especie es un linaje (clado en la terminología de De Queiroz), una secuencia genealógica de poblaciones, que evoluciona separadamente y tiene su propio rol o destino evolutivo unitario (nicho) y sus tendencias propias (libre para divergir, sin constreñimientos de otra población) (Wiley, 1978).
- El problema está en que los linajes tienen que cortarse en segmentos arbitrarios para los propósitos de la clasificación. La solución consiste en hacer la diferencia entre las especies sucesivas tan grande como la que exista entre especies contemporáneas, pero esto depende fuertemente del juicio personal.
- Otro problema es que una rama difiera del ancestro mucho menos que otra. ¿Nos encontramos ante dos o ante tres especies?
- Según la tradición cladista (Hennig 1966; Ridley 1989; Meier y Willmann 2000) cada evento de especiación entraña la extinción de la especie ancestral. Esto descarta la anagénesis, proceso mediante el que una especie se convierte en otra, sin dividirse en comunidades sincrónicas reproductivamente separadas. Desde el punto de vista de la Lógica, este modo de enfocar la especiación es correcto, pero desde le punto de vista biológico es problemático. Como criterio para la construcción de la filogenia, el cladismo utiliza la homología: Dos estructuras son homólogas si representan partes correspondientes de dos organismos construidas según el mismo plan temporal. La homología de los órganos delata la existencia de un ancestro común.
- En cada par de homólogos existen caracteres primitivos o plesiomorfias y caracteres derivados o apomorfias. La clave más importante que delata el parentesco cercano entre dos especies son las apomorfias compartidas que se denominan sinapomorfias. Los caracteres comunes de origen más remoto o simplesiomorfias no nos dicen nada sobre la cercanía del parentesco. Las autapomorfias son los caracteres derivados privativos de un taxón y carecen de significación filogenética. Un grupo monofilético incluye a todos los organismos con un ancestro común determinado.
- Ley biogenética: "la ontogenia recapitula la filogenia" (Haeckel). El desarrollo embrionario (ontogenia) permite descubrir homologías para los estados más ancestrales difíciles de observar en los individuos adultos.
- Las homoplasias tienden a confundirse con las homologías. Son estructuras semejantes, resultado de la evolución independiente a partir de estructuras ancestrales diferentes.
- En la convergencia, dos estructuras similares han evolucionado independientemente a partir de procesos de desarrollo muy diferentes.
- En el paralelismo se constatan patrones de desarrollo semejantes en líneas evolutivas diferentes.
- Las reversiones transforman una estructura "avanzada" en otra más "ancestral".
- Según el concepto filogenético de especie (Cracraft, 1983) cada especie estaría caracterizada por una combinación única de caracteres primitivos y derivados que permitiría discernirla entre todas las demás especies. Para Rosen (1979), Donoghue (1985) y Mishler (1985) una especie es el grupo monofilético menos inclusivo que puede definirse al menos por un autapomorfismo. Según este criterio las especies no se podrían ordenar en rangos fijos, como género, familia, orden, etc.
- Kevin de Queiroz (2007) equipara una especie con una metapoblación, una serie ancestro descendiente que evoluciona por separado a través del tiempo. Una metapoblación puede incluir varias subpoblaciones conectadas (Levins, 1970: Hanski y Gaggiotti, 2004). Una especie no sería por consiguiente un linaje, sino uno de los muchos segmentos que lo conforman.
- Durante el proceso de especiación, surgen en diferentes momentos las propiedades que se consideran fundamentales en otros conceptos: el aislamiento reproductivo, la distinción cuantitativa (fenética), la cualitativa (tipológica), la exclusividad de nicho, la monofilia, etc. La presencia de una sola de estas propiedades implica ya una separación de poblaciones.
Los paleoantropólogos suelen utilizar preferentemente los conceptos filogenético y evolutivo sis bien su utilización en taxones fósiles no deja de ser problemática (Groves, 2004).
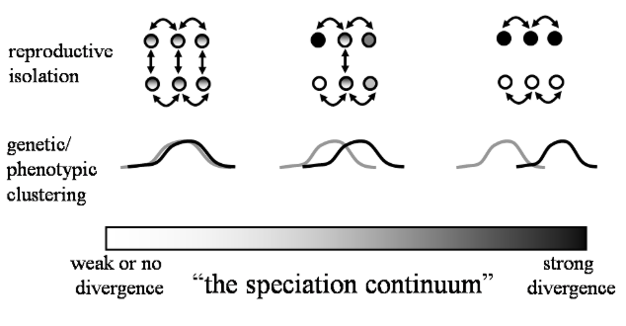 |
| El continuo de la especiación por aislamiento reproductivo. Nature Education, 2012, a partir de Nosil et al, 2009 |
Barrera reproductivas y especiación
Se distinguen varios tipos de barreras reproductivas:
- Prenupciales.
- Aislamiento ecológico o de hábitat: Los individuos potencialmente reproductores no se encuentran porque ocupan un hábitat distinto.
- Aislamiento estacional o temporal: Los individuos potencialmente reproductores no se encuentran porque tienen un periodo reproductivo distinto.
- Aislamiento sexual o etológico: Los individuos potencialmente reproductores se encuentran, pero carecen de un sistema de reconocimiento sexual.
- Aislamiento mecánico: Los individuos potencialmente reproductores se encuentran, pero tienen una estructura morfológica de los órganos sexuales que impide la cópula o la polinización.
- Postnupciales prezigóticas. Cualquier mecanismo que impide la formación del cigoto híbrido. Por ejemplo, los espermatozoides no consiguen fecundar el óvulo.
- Aislamiento gamético: En organismos de fecundación externa, los gametos de distinto sexo no se atraen mutumente y en organismos de fecundación interna, los gametos o gametofitos de una especie son inviables en el conducto sexual o en el estilo de la otra especie.
- Postnupciales postzigóticas: Los híbridos son inviables o no hallan pareja.
- Aislamiento por invibilidad híbrida: Los cigotos híbridos tienen una viabilidad reducida o nula.
- Aislamiento por fertilidad reducida o nula: Los cigotos híbridos tienen una viabilidad reducida o nula.
- Aislamiento por depresión híbrida: Los híbridos producto del cruzamiento del híbrido con una especie parental, tiene viabilidad o fertilidad reducidas.
 |
| Proceso de separación de dos especies a partir de un linaje ancestral. Las líneas paralelas indican momentos en que se adquieren propiedades como la monofilia, la distinción fenética, el aislamiento reproductivo, etc. Kevin de Queiroz (2007). |
Tipos de especiación:
Los patrones biogeográficos determinan el tipo de especiación:
- La especiación alopátrica o alopátrida tiene lugar cuando la selección natural favorece fenotipos diferentes en regiones aisladas. Aunque los grupos de individuos se junten posteriormente (contacto secundario), pueden actuar dos procesos a favor de la especiación:
- El desplazamiento de carácter puede ocurrir si la competencia por el alimento, la pareja u otros recursos incrementan las diferencias morfológicas entre los inmigrantes y los residentes.
- Si los híbridos son menos viables, la selección natural favorecerá las adaptaciones morfológicas o de comportamiento que impidan los apareamientos entre individuos de diferentes poblaciones. Este proceso es denominado refuerzo del aislamiento (Wallace; Blair, 1955; Dobzhansky).
- La Hipótesis de la dominancia en la regla de Haldane explica la inviabilidad y estirilidad híbridas debido a una interacción entre factores recesivos del cromosoma X y otros autosómicos.
- Una especie peripátrica o peripátrida ocupa un área pequeña en la periferia de la distribución de la especie de la que procede. La especiación se produce por el aislamiento de un pequeño número de individuos (demo).
- En la especiación por fundación-expansión (founder-flush) unas condiciones favorables permiten una expansión poblacional desmesurada a partir de los fundadores que desborda los recursos disponibles y colapsa. Este ciclo de fundación-expansión-colapso se repite varias veces y en cada episodio se reorganiza el genoma de un modo aleatorio favoreciendo el surgimiento de nuevas especies.
- El efecto fundador es el conjunto de cambios del genoma que se producen en los sucesos fundadores de la especiación peripátrida y de fundación-expansión.
- En la especiación vicariante, hay una división del área de distribución. Un cambio tectónico o climático aísla dos o más zonas del área de distribución de una especie, diferenciándose con el tiempo por selección gradual en cada zona aislada una población que llega a convertirse en una nueva especie.
- Fases de la especiación alopátrida:
- Fase de formación de barrera.
- Fase de divergencia por aislamiento.
- Fase de contacto secundario.
- Fase de divergencia final.
- Solo las dos primeras son necesarias para producir nuevas especies.
- La especiación parapátrica o parapátrida sostiene que puede surgir una nueva especie si la selección natural se combina con un aislamiento genético parcial (un refuerzo). Estas especies ocupan áreas de distribución contiguas.
- Se denomina clina a la variación de algún carácter de una especie siguiendo un patrón a lo largo de un gradiente latitudinal, altitudinal o cronológico. La norma de reacción es el conjunto de fenotipos (ecofenotipos o ecotipos) a que da origen un mismo genotipo cuando se desarrolla en distintos ambientes.
- Regla de Bergmann. En los vertebrados endotérmicos el tamaño aumenta progresivamente de los lugares más calientes a los más fríos. Los individuos de mayor tamaño tendrán menos superficie expuesta a la pérdida de calor que los más pequeños
- Regla de Allen. En los animales de sangre caliente, las partes sobresalientes tienden a ser más cortas en los climas más fríos ya que la radiación de calor a través de estas extremidades es desventajosa en climas frios.
- Regla de Gloger: Las poblaciones de animales de lugares calientes y húmedos, tienden a ser más oscuras que las de sitios fríos y secos.
- Efecto de Baldwin o de asimilación genética: la selección natural opera sobre las variaciones fenotipicas y estas se incorporan al patrimonio genético de la población. Una variación genotípica concede cierto grado de adaptación temporal antes de que aparezcan las mutaciones que permitan la fijación del cambio fenotipico.
- La interrupción del flujo génico se debe a la diferenciación progresiva a ambos lados de cada área transicional (ecotono), generando zonas estrechas de hibridación con selección en contra de los híbridos (zona de tensión).
- Especies anilladas: Cadenas de poblaciones que se vuelven sobre sí mismas y en los extremos del anillo se solapan sin poder cruzarse.
- Cuando una especie se encuentra ante un conjunto de nichos vacíos, puede diversificarse para llenarlos. Este proceso se denomina radiación adaptativa. Este tipo de especiación fue discutida durante mucho tiempo pero existen evidencias recientes de especiación simpátrida en peces cíclidos en lagos volcánicos de Camerún y Nicaragua (Schliewen 2001, Barluenga 2006) y de salamandras en aguas subterráneas en Tennessee (Niemiller, Miller, 2008).
- La especiación simpátrica o simpátrida tiene lugar cuando se produce una selección natural fuerte, sin necesidad de aislamiento genético. Se inicia con el establecimiento de un polimorfismo estable por selección diversificadora dentro de una población. Un genotipo seleccionado a favor en un ambiente ecológico está cada vez más aislado de los demás, llegando a constituir primero una raza ecológica y finalmente una nueva especie en simpatría.
- Se denomina contacto primario a la ocupación de un mismo territorio por dos especies incipientes que se han producido en simpatría o parapatría.
- En la especiación ecológica las barreras para el flujo de genes entre poblaciones evolucionan como resultado de la selección divergente basada en la ecología entre entornos (Rundle y Nosil 2005). La selección natural divergente actúa gracias a la presencia de distintos entornos ecológicos o favoreciendo extremos opuestos de un rasgo dentro de una única población (selección disruptiva).
 |
| La especiación es un proceso de múltiples niveles a través del tiempo y el espacio. R. Abbott et al, 2013. |
Mecanismos genéticos de especiación
Los puntos de vista sobre la naturaleza del cambio se pueden polarizar en gradualistas y saltacionistas. Según la Hipótesis de los equilibrios puntuados, (p. ej. Landis y Schraiber, 2017) los cambios evolutivos están limitados a los periodos de especiación, que se intercalan entre largos periodos de equilibrio sin cambio detectable en las especies (estasis).
- Especiación gradual (gradualismo filético): Inducida por la adaptación gradual a los cambios ambientales.
- Especiación cuántica o saltacional. Cambios que se producen por azar, pero la selección coopera a posteriori en la adaptación de las especies incipientes.
- Especiación por hibridación. Integración en un mismo genoma de características de otros por hibridación de especies.
- Especiación recombinacional. Especiación híbrida diploide que genera especies debido a la unión de gametos parentales equilibrados por recombinación.
- La especiación alopoliploide o anfiploide se inicia con la hibridación de especies y duplicación subsiguiente de los cromosomas del híbrido que permite una meiosis regular y por consiguiente niveles altos de fertilidad.
- La especiación cromosómica es debida a cambios cromosómicos que generan una o más reordenaciones que disminuyen la aptitud del heterocariotipo. La superación de esta baja aptitud se consigue mediante procesos saltacionales en una estructura poblacional microdémica.
La heterocronía como mecanismo de especiación
La velocidad de desarrollo de los individuos puede ser diferente en descendientes y antepasados (heterocronía).
- Alometría: Crecimiento relativo diferencial entre dos variables (dimensiones) del cuerpo de un organismo dependientes del tiempo de desarrollo.
- Fases juveniles de los ancestros pueden quedar retenidas durante el desarrollo en los descendientes, de forma que se eliminan las fases finales (pedomórfosis). El carácter evolucionado muestra un desarrollo juvenil respecto al antepasado.
- La neotenia produce individuos adultos con caracteres somáticos juveniles pero perfectamente maduros sexualmente.
- El posdesplazamiento consiste en un retraso en el inicio del desarrollo del carácter.
- Peramórfosis. El carácter evolucionado muestra un desarrollo extendido respecto al antepasado.
- Hipermórfosis: La evolución del carácter se produce dilatando el final del desarrollo.
- Predesplazamiento: Inicio temprano del desarrollo del carácter.
- Progénesis: Detención temprana del desarrollo del carácter.
- Aceleración: Aumento de la velocidad del desarrollo.
- Isometría: Crecimiento en el que dos variables (dimensiones) del cuerpo de un organismo cambian a la misma velocidad con el tiempo de desarrollo.
Superespecies, grados, subespecies, razas, demos
- Ernst Mayr asignó el término superespecie a un grupo de al menos dos especies en alopatría. Las superespecies consistentes en dos especies hermanas se denominan par de especies. Una especie que forma parte de una superespecie se denomina alospecie.
- Julian Huxley denominó grado evolutivo es un grupo de especies unidas por rasgos morfológicos o fisiológicos, que ha dado lugar a otro grupo que difiere notablemente de la condición ancestral, y por lo tanto no se considera parte del grupo ancestral. El grupo ancestral no será filogenéticamente completo, por lo que va a representar un taxón parafilético.
- Cuando las poblaciones muestran una diferenciación grande y el grado de aislamiento reproductivo es incipiente, se denominan semiespecies o subespecies y se dice que las especies son politípicas.
- Las poblaciones diferenciadas por la variación geográfica, se dominan razas.
- Una población mendeliana o demo es un conjunto de organismos que intercambian su material genético.
Bibliografía: Evolución: Origen, adaptación y divergencia de las especies. Antonio Fontdevila y Andrés Moya. Editorial Síntesis. 2008.
Enlaces:
Te puede interesar:
|
Enhorabuena y muchas gracias por el magnífico blog que estás realizando
ResponderEliminarFelipe