Sarah Zielinski nos resume los últimos descubrimientos sobre neandertales.
Aparición.
Para John Hawks (2012), según las evidencias
genéticas, los neandertales y la población de sapiens en África comenzaron a diferenciarse en algún momento antes
de hace 250 ka. Pero los heidelbergensis parecen
tener algunas características similares a los neandertales.
·
Una posibilidad es que los
antepasados de los africanos y los neandertales vivieran fuera de África.
·
Otra es que una población que
representa la mayor parte de los ancestros de los neandertales salieron de
África, más recientemente, tal vez en los últimos 150 ka, y se mezclan con la
población europea anterior.
·
Incluso es posible que los
ancestros neandertales y africanos vivíeran en Europa y África,
respectivamente, con una alta tasa de flujo de genes entre ellos.
Los
neandertales evolucionaron en Europa en condiciones de aislamiento geográfico y,
como consecuencia, de aislamiento genético durante todo el Pleistoceno medio. En
cambio, los fósiles del Pleistoceno medio africano (aunque muy similares morfológicamente
a sus parientes próximos del Pleistoceno medio europeo) serían antepasados exclusivamente
de Homo sapiens.
A la luz de los
hallazgos y en función de cómo se clasifiquen, tenemos varias alternativas en
cuanto al momento de su aparición:
- Una
aparición temprana, si los fósiles europeos arcaicos de los primeros
momentos del interglaciar Mindel-Riss, o incluso anteriores, pertenecen ya
al clado neandertal.
- Una
aparición intermedia, si los ejemplares de Ehrigsdorf (Weimar, Alemania),
Pontnewydd (Gales) o Saccopastore (Italia) son los primeros neandertales.
- Una
aparición tardía si entendemos con Clark Howell (1952) que los
neandertales se extienden por Europa a comienzos de la glaciación Würm
(0,115 mda).
Los neandertales constituyen un grupo
geográficamente marginal que se desarrolló en condiciones muy especiales, en el
límite del área de extensión de los homínidos, lejos de las zonas tropicales y
en una región relativamente aislada. (Hublin, 2001).
Edad.
230 ka hasta 30 ka. Un estasis considerable de 200.000 años.
Yacimientos.
Múltiples yacimientos en Europa y Asia cercana pero
no en África.
Los
fósiles de la última parte del Pleistoceno medio (desde hace unos 200.000 años)
ya pueden ser considerados, a todos los efectos, verdaderos y completos neandertales.
Algunos de los neandertales de comienzos del Pleistoceno superior (en torno a -127.000
años) son los dos cráneos de Saccopastore (Roma, Italia) y el conjunto de fósiles
de Krapina (Croacia).
A
partir de ese momento, los fósiles neandertales empiezan a ser abundantes en los
yacimientos europeos, y se encuentran también en Asia central y Oriente Próximo,
lugares a donde emigraron desde Europa. Fósiles tan emblemáticos como Le Moustier,
que da nombre al Musteriense, Guattari 1 (Monte Circeo) o La Chapelle-aux-Saints,
vivieron en Europa hace menos de 60.000 años. Estos neandertales tienen algunas
características nuevas con respecto a los anteriores, y son llamados a menudo "neandertales
clásicos". Los últimos neandertales vivieron en Europa hasta hace unos 30.000
años, e incluso algo menos, antes de extinguirse.
 |
| Área de yacimientos neandertales |
1.1.1
Anteriores a 134 ka.
En Biache-Saint-Vaast (Francia) aparecieron dos fragmentos
craneales datados entre 180-130 ka.
En la Cueva Bajondillo (Málaga) hay evidencias de
neandertales y del consumo de mariscos, con una datación de 150 ka (Miguel
Cortés-Sánchez y otros, 2011).
En Tossal de la Font (Castellón) se encontraron un fragmento
de húmero y otro de pelvis.
PRINCIPALES YACIMIENTOS
NEANDERTALES.
|
|
Iberia, Pirineos y Mediterráneo.
|
Europa Occidental
|
Europa Central, Norte y Oriental y Oriente Próximo
|
Asia
|
|
134-74 ka
|
Crombe Grenal
Saccopastore
Appidimia
Kaprina
|
|
Tabun
|
|
|
74-59 ka
|
La Quina
|
|
Subalyuk
Kebara
|
Shanidar
|
|
59-45 ka
|
El Sidrón
|
|
Khulna
|
|
|
45-35,8 ka
|
La Ferrasie
Le Moustier
La Chapelle-aux-Saints
|
|
Feldhofer
|
|
|
35,8-27 ka
|
Saint-Césaire
Gorham
|
Spy d’Orneau
|
|
|
1.1.2
Datados en el gran interglacial del periodo OIS5
(134.000-74.000 años).
Son raros y dispersos.
·
Francia
o
Cueva de Crombe Grenal,
Dordoña, Francia. Ocupada por neandertales entre 115-50 ka. Excavada en
1950-1960 por François Bordes.
§
Según el estudio de
Phillip Chase de los huesos de animales hallados en el yacimiento, los
neandertales cazaban ciervos y renos, de uno en uno, y aprovechaban la carroña
de caballos y bóvidos.
§
Indicios de utilización
simbólica del esqueleto de rapaces diurnas (Morin y Laroulandie, 2012).
o
Abrigo Bourgeois-Delaunay. Hueso temporal
primitivo.
o
Cueva Rochelot.
o
Moula-Guercy. Fragmentos craneales
o
Artenac (Charente). Fragmentos craneales.
·
Alemania
o
Taubach.
o
Salzgitter.
·
Italia.
o
Saccopastore. Un cráneo masculino y otro
femenino. Arcaicos.
·
Eslovaquia.
o
Gánovce. Fragmentos de cráneo, molde
endocraneano y otros de poscraneanos.
·
Grecia.
o
Acantilado de Apidima.
§
Apidima I. Medio cráneo.
§
Apidima II. Varios fósiles humanos con partes
del cráneo facial. Primitivos.
·
Croacia.
o
Kaprina. Cientos de fósiles de unos ochenta
individuos. Primitivos. Se infieren prácticas caníbales. Descubierto
por Dragutin Gorjanovic-Kramberger en 1899 cerca de Krapina (Croacia). este
yacimiento proporcionó restos de dientes y fragmentos de mandíbuulas. Cuando
Gorjanovic publicó sus hallazgos en 1906, descartó que los neandertales fueran
una patología de los humanos modernos.
·
Península Ibérica.
o
Lezetxiki, Mondragón, Guipúzcoa. Primer molar y
segundo premolar.
o
Bolomor, Tavernes, Valencia. 5 dientes
y fragmento de fémur.
o
Tossal de la Font, Castellón. Dos
restos poscraneales.
o
Cova Negra de Bellús, Xátiva, Valencia.
Nivel 33. Fragmento mandibular.
o
Pinilla del Valle, Madrid. Dos dientes.
o
Los Casares, Riba de Saelices,
Guadalajara. Metacarpiano. Extraviado.
o
Cueva de Valdegoba, Huérmeces del
Cerro, Burgos. Mandíbula, varios dientes, falange de la mano, dos metatarsianos
y otros, pertenecientes al menos a cinco individuos distintos.
o
Carihuela, Granada. Carihuela-3. Hueso
frontal de un niño, depositado por algún depredador.
o
Cova Eirós, Triacastela, Lugo.
Artefactos que fueron utilizados como armas de caza. Otros útiles se emplearon
para despiece. Otra parte del utillaje sirvió para cortar y tallar madera,
posiblemente para fabricar los mangos de las armas de caza. Por último, otros
artefactos se usaron para preparar pieles secas que supuestamente sirvieron
para confeccionar vestimentas.
1.1.3
Datados en la glaciación del periodo OIS4
(74.000-59.000).
·
Francia.
o
Regourdou. Sepultura datada hace 70.000 años.
o
La Quina. Fósiles humanos de al menos 27
individuos, con industria asociada.
§
H5. Esqueleto incompleto de adulto.
§
H9. Mandíbula de adulto.
§
H18. Cráneo infantil.
·
Hungría.
o
Subalyuk. Esqueleto poscraneal, mandíbula de
adulto y cráneo infantil, con industria musteriense.
·
Chequia.
o
Ochoz. Mandíbula de adulto, molar y fragmentos
de huesos poscraneales.
·
Italia.
o
Caverne delle Fate, Savona, Liguria. Frontal,
occipital, zigomático, fragmentos de mandíbulas y dientes sueltos.
·
"Shanidar Site". Ralph Solecki
descubrió 9 esqueletos de neandertal entre 1953 y 1960 en la cueva de Shanidar (Irak).
Su edad está comprendida entre los 70 y 40.000 años. Uno de ellos, Shanidar 4, fué
enterrado con una ofrenda de flores (aunque algunos lo discuten) En 1971 Solecki
escribió el libro "Shanidar, the First Flower People", en el que trataba
de quitar los estereotipos de semi-humanos brutos acuñado para los neandertales.
Otro esqueleto, Shanidar 1, estaba parcialmente ciego y mutilado de un brazo; si
sobrevivió fué porque le ayudaron, lo cual demuestra una compleja organización social.
·
Península Ibérica.
o
Covalejos, Cantabria. Dos dientes de muda.
o
Cova Negra de Bellús, Xátiva, Valencia.
Niveles 20-17. Restos de 7 individuos (4 niños, 1 adolescente y 1 adulto). Cráneo
4, radio y fragmento de peroné, con un hogar a techo.
1.1.4
Datados en el tramo templado OIS 3c
(59.000-45.000 años).
·
Cueva Malarnaud (Francia). Mandíbula de adulto.
·
Khulna (Chequia). Cráneos, mandíbula y
poscraneales de al menos 11 individuos. Maxilar derecho, pragmento de parietal
derecho y dientes aislados datados en 45.600 años.
·
Carihuela-2. Nivel VI.
·
Sima de las Palomas, Cabezo Gordo, Torre
Pacheco, Murcia. 89 restos, entre ellos diversos fragmentos craneales y mandibulares.
·
Cueva del Gegant de Sitges, Barcelona.
Mandíbula.
·
La Güelga, sector oriental del Macizo
Asturiano.
·
El Sidrón, Piloña, Asturias. Más de
2.000 fósiles (213 de dientes) de al menos 13 individuos: uno infantil (2-3
años) otro infantil casi completo (6-7 años, hijo de una de las mujeres del
grupo y hermano del anterior) con molares de leche e incisivos definitivos
incipientes), otro juvenil, dos de adolescentes, siete de jóvenes adultos (tres
hembras, tres machos y uno indefinido). Asociados a 415 artefactos líticos y 51
restos faunísticos.
o
Se infiere que yacían enterrados próximos
a una entrada de la cueva y fueron arrastrados con el sedimento que les
envolvía. Pueden ser considerados miembros de una misma población o grupo
familiar.
o
Datados en 43 ka.
o
Dientes de tamaño grande. Los incisivos
en pala, muy curvados y con fuertes tubérculos en los bordes laterales
internos. Los premolares, asimétricos, con cúspides accesorias y crestas
transversas. Algunos molares taurodontos.
o
Tres mandíbulas están completas o casi.
El cuerpo mandibular es alto y grueso. Trígono mentoniano, sin escotadura.
Espacio retromolar corto comparado con otros neandertales.
o
La comparación con otros neandertales
sugiere variabilidad racial y geográfica.
o
Cálculos en los dientes de todos los
adolescentes y adultos; absceso apical en la mandíbula 2, indicador de
periodontitis; en la mitad de las piezas dentarias se observan surcos en
sentido vertical en la cara anterior de un diente y la posterior del contiguo,
de etiología problemática; hipoplasia del esmalte dentario que señala
deficiencia nutricional; sarro; descascarillado.
o
Los huesos presentan marcas y cortes de origen
antrópico, las de los huesos largos como para desarticularlos rompiendo
ligamentos. Fractuas indicativas de extracción de médula y sesos. Parece fuera
de duda la práctica de canibalismo alimentario.
o
En una adulta joven, se han hallado restos de
hierbas medicinales amargas: manzanilla y milenrama, lo que sugiere
automedicación gracias al conocimiento de las cualidades medicinales.
1.1.5
Datados en el intervalo frío OIS 3b
(45.000-35.800 años)
·
Francia.
o
La Ferrasie. Restos de dos esqueletos adultos y
unos cinco infantiles, datados en 45.000 años.
o
Le Moustier.
§
Esqueleto infantil (extraviado) datado en 42.500
años
§
Esqueleto de subadulto, datado en 40.900 años.
o
L’Hortus, Languedoc. Restos de unos 20 individuos
datados en 40.000 años.
o
La Chapelle-aux-Saints.
Old Man, descubierto
por Amédée Jean Bouyssonie en 1908. tiene una antigüedad de 40.000 años y una
capacidad craneal de 1.620 c.c. El esqueleto casi completo fué reconstruido por
Marcellin Boule quien exageró los ragos anatómicos, popularizando el
estereotípo de hombre mono. Se confundió porque no se dió cuenta que el
individuo muerto a los 30 o 40 años tenía una artítris avanzada que
distorsionaba sus rasgos y doblaba sus hombros.
·
Italia.
o
Cueva de Fumane. Evidencias de obtención de
plumas de aves para uso simbólico.
·
Chequia.
o
Dzeeraba Skala. Molar inferior. Datado con dudas
en 40 ka años.
·
Bulgaria.
o
Bacho Kiro. Datado en 43 ka.
·
Croacia.
o
Vindija. Cráneos, mandíbula y poscraneales.
Datado en 42 ka. Resultado de prácticas caníbales.
·
Alemania.
o
Feldhofer, Neander. Pequeña cueva. En
1856 Johann Fuhlrott halló una bóveda craneal, huesos del fémur, parte de una
pelvis, algunas costillas y huesos del brazo y hombros. El individuo al que
pertenecían se rompió el antebrazo izquierdo en vida, dando como resultado que
los huesos del brazo izquierdo son más cortos que el derecho.
·
Península Ibérica.
o
Carihuela-1. Nivel 5.
o
Arrillor. Muela infantil.
o
Axlor. Maxilar incompleto y fragmento
de mandíbula con varios dientes, de un mismo individuo.
o
El Castillo, Puente Viesgo, Cantabria.
Más de 30 fósiles de unos doce individuos.
o
Banyoles. Mandíbula de adulto avanzado,
con dientes muy gastados.
o
Oliveira, Almonda. 4 fragmentos.
o
Cova Negra de Bellús, Xátiva, Valencia.
Niveles 13-11. 2 metatarsianos, fragmento de diáfasis femoral, parietal-2.
o
Calvero de la Higuera (Madrid) Lozoya, dos incisivos, un canino y una
muela de una menor de 2 años y medio (2011), datados en más de 40 ka. Hay
indicios de que el cuerpo de la niña fue depositado intencionalmente.
1.1.6
Datados en el intervalo oscilante OIS 3a
(35.800-27.000)
·
Francia.
o
Arcy-sur-Cure. Datado en 33.800 años.
o
Saint-Césaire. Esqueleto aplastado descubierto
por Francois Leveque en 1979. El cráneo está casi completo, faltándole la parte
de abajo. Se ha datado alrededor de 35.000 años, y es uno de los últimos neandertales
conocidos. Este yacimiento es importante porque se encontraron herramientas que
fueron copiadas a los primeros cromañones.
·
Bélgica.
o Gruta de
Spy d' Orneau. Cráneos, mandíbulas y poscraneales (dos esqueletos casi completos
Spy 1 y Spy 2) descubiertos por Marcel de Puydt y Max Lohest en 1886. Datados
entre 38.000-34.000 años. El hallazgo sirvió para desacreditar la idea de que los
neandertales sufrían una especie de disminución psíquica.
·
Italia
o
Cavallo. Datada en 33.000 años.
·
Península Ibérica.
o
Cueva de Gorham,
Gibraltar. Forma parte de un conjunto de cuevas ocupadas por neandertales.
Desde hace 125 ka, el clima fue similar al actual. Los neandertales explotaban
un hábitat similar al actual de Doñana, con abundancia de presas pero también
con depredadores como leones, linces, lobos y osos pardos. La costa se hallaba
a unos 5 km. de la cueva. Los neandertales comían grandes mamíferos, con
preferencia por la cabra montés, pero los restos más abundantes corresponden a
conejos; también son muy abundantes los restos de aves. Comían también tortugas
de tierra, piñones, lapas, mejillones, focas y delfines. Es de suponer que
comiesen también frutas, raíces y larvas, que no dejaron rastro. Datados en
30-26 ka aparecen restos auriñacienses, lo que puede indicar la convivencia con
sapiens.
o
Devil’s Tower. Media
docena de fósiles incluyendo un cráneo infantil icompleto y una mandíbula.
o
Covalejos. Nivel 8.
o
El Salt, Alcoi,
Alicante. 6 dientes.
o
Cueva de los Moros de
Gabasa, Ribagorza, Huesca. Premolar, dos molares, metatarsiano, falange del pie,
clavícula.
o
Columbeira. Germen de
primer molar.
o
Boquete de Zafarraya,
Alcaucín, Málaga. 3 mandíbulas, escápula, húmero, pubis, 3 fémures, tibia y
diente lacteal de nueve individuos.
o
Figueira Brava,
Sesimbra. Nivel C-2. Premolar superior.
o
Salemas, Ponte de Lusa.
Infantil: varios huesos y un maxilar con dientes y una muela de leche.
o
Cova Negra de Bellús, Xátiva, Valencia.
Niveles 9 y 8. Cráneos 3 y 5.
o
Cova Foradá de Oliva. Valencia. Roca
con forma de cabeza de oso cavernario hallada junto a restos humanos, con
incisiones.
1.1.7
Últimos neandertales.
En 2011 Javier Baena et al comunicaron las dataciones de los
hallazgos en la Cueva del Esquilleu, Liébana, Cantabria, España. Los restos más
modernos ofrecen una datación de 20 ka. La tecnología lítica asociada corresponde
al último musteriense.
Características.
Siguiendo a Clark Howell
(1952) los rasgos morfológicos de los neandertales clásicos son los siguientes:
- Menos platicefalia que los erectus,
pero mantenida hasta cierto punto. Depresión central en el occipucio (fosa
suprainíaca). La parte posterior del cráneo tiene un abultamiento
característico (moño) que no está concentrado en un punto, como en el caso
del erectus; servía para anclar
una fuerte musculatura del cuello.
- Esqueleto facial ancho con nariz prominente y gran arco cigomático.
- Gruesos arcos supraorbitales semicirculares y separados que no se
extienden lateralmente. A diferencia de heidelbergensis y rhodesiensis
estas estructuras óseas están bien desarrolladas en las hembras y
aparecen en una fase temprana del crecimiento, hacia los ocho años. Disponen
de senos frontales interiores que aligeran esa estructura en apariencia
tan masiva. En los erectus, son
compactos.
- Ausencia de barbilla. Mandíbula potente, con un diastema retromolar
entre el M3 y el ramus de la mandíbula. Incisivos relativamente grandes,
que muestran un fuerte desgaste. Molares de tamaño reducido. Taurodontismo
molar: no ramificación de las raíces y grandes cavidades pulpares.
- Trinkaus (1988) lo considera como el resultado de la combinación
de dos factores:
i. El mantenimiento
del prognatismo facial del grado erectus (plesiomorfia)
ii. La retirada hacia
atrás de la región masticatoria muscular (sinapomorfia compartida con los sapiens).
- Según Loring Brace (revolución culinaria) las características
faciales guardan una relación estrecha con la importancia de los incisivos
para la manipulación de objetos y la fabricación de herramientas. La
dirección de las marcas sugiere que la mayoría de los neandertales eran
diestros. A medida que la industria lítica mejora, comienza a declinar el
tamaño de los dientes y el rostro.
- Las más grandes capacidades craneales en un homínido (media de 1.500
cc).
- Trinkaus (1984) sugiere que puede ser el resultado de un periodo
de gestación más extenso.
- Poirier (1987) las atribuye a un cuerpo más pesado.
- Esqueleto postcraneal muy robusto. Columna vertebral corta y
masiva. Extremidades robustas y cortas en relación a la altura.
Articulaciones grandes. Omóplatos con inserciones musculares extensas.
Caja torácica grande y redondeada.
- Las extremidades cortas serían para Trinkaus un rasgo derivado,
pero entran dentro del rango de variabilidad de los humanos actuales, por
lo que no cabría considerarlo como un rasgo propio.
- De los estudios genéticos realizados con los neandertales de El
Sidrón, se deduce que eran de piel clara, con el color del pelo rojo parecido
al de los caucásicos (variante del gen MICR). Poseían una versión similar
a la nuestra del gen Foxp2, relacionado con el lenguaje. (Finlayson,
2010).
Trinkaus (1988) solo acepta como verdaderas
apomorfias de los neandertales unos pocos rasgos relativos a las regiones
temporales y occisito-mastoideas del cráneo. Esta convicción no es ajena a la
idea los neandertales como una variedad de nuestra propia especie.
Schwartz y Tattersall (1996) consideran algunos
rasgos en la región interna de la nariz como autapomorfias y por tanto están a
favor de la separación de especies.
Lieberman y McCarthy (1999) opinan a favor de la
separación de especies, basándose en la falta de proyección facial de los
humanos modernos.
Conviene advertir que estos rasgos no están
presentes en todos los ejemplares. Existe un rango muy amplio de variedad
atribuida por Wolpoff (1980) en gran medida a los dimorfismos sexuales.
Diferencias y semejanzas entre sapiens modernos y neandertales.
1.1.8
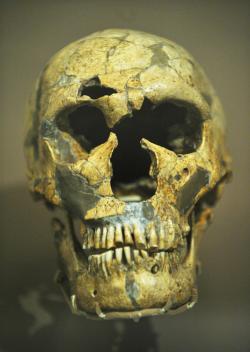
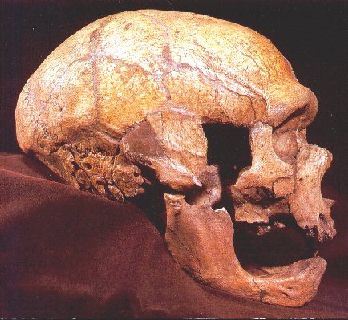
El cráneo.
La caja craneana de los neandertales es
muy grande. Vista de perfil es larga y baja, con la frente huida; el occipital,
alargado y proyectado hacia atrás, pero redondeado, tras una depresión hacia
arriba (fosa suprainíaca). Los cráneos de los neandertales tienen en vista posterior
un perfil redondeado mientras que nuestro cráneo visto por detrás tiene un perfil
pentagonal.
En los neandertales faltan las fosas
parietales latero-súperas y la anchura máxima de la bóveda se mantiene baja,
entre los temporales mientras que en los modernos, la altura máxima se eleva
hasta los parietales.
El cráneo de los neandertales presenta un torus
supraorbitario muy marcado, grueso y saliente, principalmente en el centro,
arqueado y debilitado en la gabela, entre las órbitas, y lateralmente forma dos
arcos que continúan el contorno de los ojos y contienen grandes senos frontales.
La cara es muy prognata, la frente es baja.
Los neandertales poseían grandes capacidades
cerebrales (entre 1.200-1.700 cc, volúmenes similares a los de nuestra especie e
incluso ligeramente mayores). Los neandertales y nuestra especie somos las dos formas
humanas más encefalizadas, pero el crecimiento del tamaño del cerebro se produce
de forma independiente en ambas líneas evolutivas.
-
el neurocráneo del
hombre moderno es alto y de frente levantada.
-
el cráneo de los
neandertales se hizo muy alargado.
Markus Bastir, Chris Stringer et al
(2011), tras comparar los cráneos, han sacado tres conclusiones:
1. Los lóbulos temporales en sapiens parecen haber
crecido mucho más a lo largo de la evolución de nuestra especie. Esto ya se
sabía cuando se comparaba con chimpancés, por ejemplo, pero hasta ahora no se
había podido saber si era una característica de todos nuestros antecesores o
si, como parece haber confirmado esta investigación, ese tamanaño inusualmente
grande los lóbulos temporales es propio solo de nuestra especie. El lóbulo
temporal es una parte del cerebro, localizada frente al lóbulo occipital,
aproximadamente detrás de cada sien, que desempeña un papel importante en
tareas visuales complejas, como el reconocimiento de caras. También recibe y
procesa información de los oídos, contribuye al equilibrio, y regula emociones
y motivaciones como la ansiedad, el placer y la ira.
2.
El lóbulo
olfativo también es más grande que los de los neandertales (en tamaño absoluto
y relativo).
3. La forma de los lóbulos frontales es distinta en sapiens y neanderthalensis. En el lóbulo frontal se encuentra el área de
Broca, encargada de la producción lingüística y oral y se regulan los
movimientos de los órganos fonoarticulatorios.
La cara de los neandertales tiene prognatismo medio-facial,
es decir, está proyectada hacia delante. Las órbitas oculares eran grandes y
redondas. Al tener la cavidad nasal más adelantada los huesos nasales se disponen
casi horizontalmente. En los neandertales la placa ósea de debajo de las órbitas
se dispone diagonalmente (en el hombre moderno la placa infraorbitaria tiene una
orientación coronal), con lo que la cara de los neandertales presenta una apariencia
apuntada en forma de cuña, que le da un aspecto "aerodinámico". También
tienen una abertura nasal muy ancha, y la cavidad interna también es muy grande,
por ese motivo se piensa que sus narices serían planas y anchas. Se ha planteado
la hipótesis que los neandertales tenían cavidades nasales anchas para calentar
y humedecer, durante la respiración, el aire frío y seco que habría en épocas glaciares
en Europa. Sin embargo, en poblaciones actuales, no existe ninguna relación entre
el tamaño de la abertura y cavidad nasal y el clima. Parece más probable que los
neandertales necesitaban tener un gran tamaño de nariz por la necesidad de
oxigenar su cuerpo tan robusto. Las grandes fosas nasales podían
proporcionarles un fuerte vozarrón.
La anatomía del oído revela que los neandertales
tenían una capacidad auditiva similar a la de los humanos actuales. Este hecho,
junto a la también parecida configuración del gen FOXP2, favorece la teoría del
habla neandertal.
Los arcos dentarios estaban desplazados de debajo de
la base del cráneo hacia delante. Los dientes de los neandertales son del mismo
tamaño que los de nuestra especie, pero en relación al cráneo, a la mandíbula y
al tamaño corporal son relativamente pequeños. La relación entre la dentición anterior
(incisivos y caninos) y la dentición posterior (premolares y molares) es diferente:
los neandertales tienen la dentición anterior relativamente más grande. Los
incisivos superiores, en pala. Los molares tienen la cavidad pulpar expandida (característica
que se denomina taurodontismo). Además, la dentición anterior de la mayor parte
de neandertales está muy desgastada, porque utilizaban los dientes con una función
especial (quizás para curtir las pieles, o para sujetar el alimento u otros objetos
con los dientes).
La mandíbula carece de mentón, tiene el foramen mandibular
situado a la altura del primer molar y posee un espacio entre el último molar y
la rama mandibular, que se denomina espacio retromolar Este espacio aparece porque
la dentición tiene un reducido tamaño y está en una situación adelantada con relación
al hueso.
1.1.9 Forma general
En las primeras interpretaciones de la forma corporal de
los neandertales se les atribuía un aspecto brutal y simiesco. Sin embargo, hoy
sabemos que su postura erguida y su forma de locomoción sería muy similar a la de
la humanidad actual.
El promedio de estatura masculina se encuentra en torno
a 170 cm y el promedio femenino alrededor de 160 cm. Pero su complexión física es
extraordinariamente robusta, su esqueleto es mucho más corpulento, el grosor de
los huesos es mayor y las inserciones musculares están más marcadas. Los neandertales
también tienen el tamaño de las articulaciones relativamente más grandes.
Las estimaciones del peso de algunos individuos masculinos
neandertales arrojan cifras de alrededor de 80-90 kg, bastante superiores a las
de los hombres modernos de la misma estatura. Pero es probable que los pesos de
los neandertales estén infraestimados, porque las rectas de regresión utilizadas
para establecer el peso, a partir de medidas esqueléticas, se basan en estudios
sobre poblaciones actuales que tienen un cuerpo más grácil. Tampoco se tiene en
cuenta el peso más elevado que tendrían los huesos de estos homínidos, que presentan
un mayor grosor en sus paredes óseas.
1.1.10 Huesos poscraneales.
Los huesos de los neandertales eran robustos y
pesados, con un periostio grueso.
Las extremidades de los neandertales son relativamente
más cortas, especialmente en los segmentos distales, es decir huesos del antebrazo
(cúbito y radio) y pantorrilla (tibia y peroné).
Los neandertales reflejan en su forma corporal (bajos,
corpulentos y con extremidades relativamente cortas) adaptaciones al clima
frío. Para Ryan Higgins (2011), las piernas cortas son más eficientes en
ambientes montañosos.
En el coxal, los neandertales tienen el hueso púbico (más
concretamente su rama horizontal) muy alargado y aplanado. También Australopithecus
tiene el pubis largo; así, esta morfología parece ser una característica primitiva
que presentan todos los homínidos, a excepción de los humanos modernos, los únicos
que poseemos un pubis acortado y grueso. A partir de un antepasado primitivo de
gran complexión física con caderas anchas y cerebro reducido, evolucionaron dos
líneas independientes en Europa y África. La primera de las líneas evolutivas conduciría
a los neandertales y la segunda a los humanos modernos. En ambas líneas se produjo
un aumento del tamaño cerebral. Pero, mientras que los neandertales conservaron
el modelo corporal de complexión robusta y cuerpos anchos de sus antepasados, reduciendo
la longitud de sus extremidades, los humanos modernos redujeron la anchura de las
caderas y del tronco, y disminuyó su peso corporal.
En los neandertales, se aprecia un surco dorsal en la
escápula. La forma de esta y la clavícula larga evidencian una espalda ancha en
los hombros.
1.1.11 Rasgos culturales.
El conjunto de útiles líticos de los neandertales se conoce
como cultura Musteriense, y se encuadra dentro del Modo Técnico 3 o Paleolítico
Medio. El Musteriense no es exclusivo de los neandertales y en las cuevas de Qafzeh
y Skhul (ambas en Israel) hemos encontrado niveles con industria Musteriense asociados
a enterramientos de Homo sapiens.
En 2011 el equipo de Eudald Carbonell encontró las
huellas hasta el momento más antiguas de un artefacto de madera con mango. Se
trata de un negativo sobre travertino de un utensilio parcialmente carbonizado,
hallado en el yacimiento del Abric Romaní, en Capellades (Barcelona), datado en
56 ka.
En 2012 se publicó una datación de más de 42 ka para
unas pinturas rupestres en la Cueva de Nerja, pero la datación es indirecta y
falta confirmación.
Muchos de los esqueletos neandertales excavados en cuevas
han sido históricamente considerados como el resultado de prácticas funerarias.
Existen mayores dudas sobre la existencia de un ritual acompañante, es decir, si
el enterramiento tenía algún tipo de significado simbólico. Las pruebas de ritualidad
en los enterramientos neandertales son muy dudosas
Son los neandertales los que generalizan el uso del fuego
en todos sus asentamientos. En los yacimientos del Paleolítico medio aparecen frecuentemente
hogares muy bien estructurados, con los que los neandertales organizaban el espacio
donde vivían. Estos fuegos eran mantenidos quemando madera o huesos.
También tenemos pruebas de que los neandertales proporcionaban
algún tipo de ayuda social a individuos muy viejos o con incapacidades físicas
importantes.
En el yacimiento croata de Kaprina también encontramos
evidencias de canibalismo entre los neandertales: los huesos están fracturados intencionalmente,
muchos de ellos presentan marcas de corte en su superficie producidas por instrumentos
líticos, y algunos de ellos están quemados.
A partir de los conjuntos de restos de animales encontrados
en los diferentes yacimientos del Paleolítico medio, sabemos que los neandertales
eran buenos cazadores, aunque quizás algunas de sus presas también las obtuviesen
practicando métodos oportunistas y de carroñeo. Sus presas principales eran los
ciervos, caballos, bisontes, cabras, renos y, en menor medida, mamuts y rinocerontes.
En algunos yacimientos la mayor proporción de alguno de estos taxones sobre los
demás no refleja las preferencias cinegéticas de los neandertales, sino que refleja
la abundancia de este animal en el entorno inmediato de los yacimientos. Los
neandertales usaban lanzas, pero empujándolas, no lanzándolas como los sapiens. La capacidad de lanzar objetos requiere
de una coordinación extremadamente buena y de habilidades motoras, que son
proporcionadas por dos estructuras cerebrales excepcionalmente desarrolladas en
los sapiens: el cerebelo y la corteza
parietal posterior. Estas regiones estaban menos desarrolladas en los neanderthalensis.
Además de por su alimento, algunos de los animales
cazados eran apreciados también por su piel, huesos y tendones, usados para hacer
ropa, tiendas, etc. La evidencia de que usaban ropas es indirecta. El estudio del
desgaste que tienen sus útiles líticos nos indica que los usaban para cortar pieles
y darles forma.
Modos de vida.
La morfología de los neandertales clásicos se ha
considerado como el reflejo a una adaptación a un clima frío:
v Clark Howell
(1952) sostiene que las condiciones de las áreas periglaciares tuvieron un
efecto marcado en la morfología y distribución de los neandertales. En los
meses de verano abandonaban los lugares protegidos para cazar. Una migración
los llevó desde Italia y por el Norte del Adriático hacia el corredor de Europa
Central.
v (Brose y Wolpoff,
1971): orificio nasal muy ancho, con mucosa ciliar distinta y cavidad sinusal
muy voluminosa para calentar y humedecer el aire.
o
Heim (1997) disiente, señalando que existen
neandertales en lugares de clima cálido y húmedo.
v Trinkaus (1981)
está de acuerdo en que la forma de los huesos de las extremidades inferiores
expresa una adaptación a un clima frío, pero recuerda que este rasgo puede
también interpretarse por motivos biomecánicos (fuerza a expensas de velocidad
de carrera).
El desgaste de los dientes es importante y
habitual. Ha sido entendido como una utilización de la dentadura a guisa de
herramienta, pero esta hipótesis es muy debatida.
- Viven en cuevas cerca de ríos, pero no construían refugios.
- En Molodova, al este de Ucrania, se hallaron restos de una
construcción realizada con huesos de mamut y decorada con pinturas y
relieves en las paredes. La estructura está compuesta de 116 grandes
huesos incluyendo cráneos de mamut, mandíbulas y 14 colmillos y fémures.
El habitáculo, que tiene hasta ocho metros en sus partes más anchas, fue
construido con los huesos de un animal que probablemente cazaron los
mismos neandertales y se le calcula una antigüedad de unos 44.000 años. Tenía
probablemente una techumbre de madera o algún otro material ligero y era
utilizada como vivienda habitual, pues se han descubierto restos de hasta
25 hogueras en su interior. Se trata de la construcción más antigua
documentada en este tipo de materiales.
- El territorio de acción raramente superaba los 1.000 km2.
- Dominio del fuego. (200.000 años).
- Recolectores, cazadores y carroñeros. Lanzas de madera que se
empujaban. Hallazgos en Mauran (Pirineos franceses) y La Cotte (Jersey)
indican caza masiva de grandes presas por emboscada o despeñamiento. Caníbales,
todo indica que en épocas de escasez alimentaria se daba muerte a los
miembros de grupos ajenos para su consumo. Un estudio de los restos del TD10-1
de Gran Dolina en Atapuerca, con una antigüedad aproximada de 300 ka, y de
los niveles XVII, XI y IV de la Cova del Bolomor ha revelado la misma
variedad y oportunismo en el consumo de recursos alimenticios, variedad de
estrategias y diversidad comportamental que luego caracterizará al sapiens (Ruth Blasco, 2012). En el
yacimiento de Preresa (Madrid) se hallaron huesos de elefante con marcas
de corte y percusión para extracción de la médula ósea.
- Realizaban trabajos artesanales cuyo conocimiento era transmitido
entre generaciones. Sin embargo, eran poco innovadores y utilizadores de
símbolos.
- Cultura lítica musteriense (casi exclusiva de Homo neanderthalensis) (yacimiento de Le Moustier): Herramientas
de piedra que produce lascas con filos por la técnica de Levallois (golpear
suavemente u núcleo alrededor de su perímetro). Raederas y puntas de flecha.
En ambientes cambiantes, surge la cultura chatelperroniense. Desconocían
la aguja con ojo.
- Se han hallado herramientas musterienses en islas griegas y Creta.
Son posibles las siguientes explicaciones.
- Las actuales islas estaban por entonces unidas al continente. Esta
posibilidad ha sido descartada por George Ferentinos (2012).
- Los neandertales nadaban. Esta explicación puede valer para
algunas islas, pero no para Creta, distante 40 km de la tierra más
cercana.
- Los neandertales practicaban la navegación. No hay constancia de
ello, pero si admitimos la navegación para Homo floresiensis con más razón la podemos considerar para neanderthalensis.
- Debido a su gran esqueleto necesitarían mucho calcio, sólo conseguido
con una rica dieta vegetal. Sin embargo, en latitudes septentrionales,
eran casi estrictamente carnívoros.
- Morían jóvenes. Ninguno vivió más de 45 años. La esperanza de vida
media era de 35 años. Cuidados a dependientes y empatía, como muestran El viejo de La Chapelle-aux-Saints y
Shanidar-1. Katherine Dettwyler se muestra escéptica. Por otra parte, hay
pocos hallazgos de individuos impedidos de caminar, lo que podría indicar
que eran abandonados.
- Patrilocalidad, según lo que se desprende del estudio genético de
12 individuos hallados en la Cueva del Sidrón (España), victimas de un
desprendimiento, datados en 49.000 años. Vivían en grupos pequeños, de
unos 10 individuos, y aunque alguna vez se relacionaban con otras
comunidades, no hay constancia de actividad comercial ni intercambios a
grandes distancias. Existían las relaciones de pareja y de familia.
- Enterramiento de sus muertos (capacidad cognitiva y simbólica) (El viejo
de La -Chapelle-aux-Saints). Dudosas pruebas de rituales funerarios.
- Perforaciones y grabados en huesos y piedras (críticas achacándolas
a acciones naturales).
- Conducta artística (los grabados carecen de utilidad y siguen una disposición
geométrica con intención estética). En 2012 Mark J. Sier et al comunicaron
el análisis de unas manchas rojas halladas entre 1980-1983 en
Maastricht-Bélvèdere, datadas en 250 ka y que resultaron ser pigmentos de
ocre fabricados con un mineral que trasladaron al menos desde 60 km; las
manchas debieron de ser el resultado de pérdidas accidentales de material
mientras era utiizado. El problema de los restos artísticos es su tremenda
escasez en comparación con la explosión artística del Paleolítico Superior.
Según Clive Finlayson et al (2012), los neandertales utilizaban plumas de
aves como adornos personales.
- Lenguaje muy rudimentario.
Relaciones entre los neandertales y los humanos modernos.
Los Homo sapiens sapiens conviven con Homo neanderthalensis en Eurasia La discusión sobre las relaciones filogenéticas
entre neandertales y seres humanos de aspecto moderno se traduce en tres posturas
encontradas:
- Los neandertales son una especie anterior a Homo sapiens y ancestral
de ésta. Es una idea de principios del siglo XX a la que se ha adherido Wolpoff
si bien considera que forman una única especie.
- La presencia de restos neandertales clásicos en época tardía es un
argumento en contra. Podría aducirse que el salto evolutivo se hizo de forma
local – en Próximo Oriente- mientras que en otras partes se mantenía la
especie neandertal.
- Los neandertales son una subespecie del Homo sapiens, Homo sapiens
neanderthalensis (B. G. Campbell, 1963) que, contribuyó con su acervo genético
a la aparición del Homo sapiens sapiens (hipótesis de la evolución multirregional).
- Para Trinkaus, de acuerdo con las evidencias fósiles de Próximo Oriente,
sería más que probable. El estudio de las evidencias morfológicas del esqueleto
casi completo de un niño neandertal, del yacimiento del Abrigo do Lagar Velho
(Portugal), de 24.500 años, llevado a cabo por Trinkaus, indica evolución
en mosaico. Algunos rasgos, como la mandíbula incipiente o el tamaño de las
falanges del pulgar, muestran una cercanía respecto a los rasgos tenidos por
derivados de los seres humanos de aspecto moderno, mientras que otros, como
el tamaño de los incisivos o las proporciones del fémur, están dentro de los
propios de los neandertales. Ante esta situación intermedia, con rasgos en
diferente grado de evolución, concluye que, al menos en la península Ibérica,
se produjo un cruce de las poblaciones. Considera además que los rasgos de
conducta entre neandertales y humanos estaban lo bastante próximos entre sí
como para permitir el intercambio genético.
- Los estudios genéticos han revelado que las poblaciones de sapiens
actuales, salvo las del África subsahariana, comparten variaciones de
genes con los neandertales. Esto podría tener varias explicaciones:
i. Los humanos
modernos absorbieron genes neandertales en el Levante, y luego se distribuyeron
uniformemente a través de Eurasia.
ii. En la población
africana se produjo la separación del antepasado de neandertales y humanos
modernos. En África, los humanos modernos se mantuvieron alejados de los
neandertales mientras que en Eurasia se dieron mezclas. (Estructura africana
profunda de la población + antepasado neandertal-sapiens común en África del
Este)
iii. Los hallazgos
paleontológicos y arqueológicos sugieren que los seres humanos modernos y los
neandertales coincidieron en la región mediterránea del este hace alrededor 100
mil años durante una época en que la fauna africana alcanzó temporalmente el
Oriente Medio. Los sapiens modernos se contrajeron luego hacia África,
separándose de los neandertales. Hace 50.000 años, las poblaciones del este de
África protagonizaron la última salida, empujados por las poblaciones del
centro y sur de África. Los genes neandertales pudieron haberse fijado en la
población que salió de África, gracias al efecto fundador.
1.
El descubrimiento de seres humanos modernos en el
sur de China, datados en 0,1 mda hace menos posible esta alternativa. Cualquier
idea del retratamiento llega a ser inverosímil.Son dos especies diferentes con
los neandertales formando una rama lateral que no guarda relaciones de ancestro
con Homo sapiens. De hecho éste
podría ser incluso más antiguo. Es la postura de Stringer y Gamble.
(Sustitución brusca de las poblaciones anteriores de neandertales por seres
humanos de aspecto moderno. Hipótesis del reemplazo).





















{kind=link}